辅酶Q10
辅酶Q10
辅酶Q10是一种脂溶性醌类化合物, 又名辅酶q10、癸烯酸和维生素Q10, 它作为人体中唯一的辅酶Q类物质, 参与线粒体氧化磷酸化与ATP的产生过程, 调控细胞氧化还原反应环境, 在电子穿膜过程中携带还原电子进入囊泡或带出胞外, 参与内膜和质膜的质子梯度形成, 是细胞呼吸和代谢的激活剂, 也是重要的抗氧化剂和非特异性免疫增强剂。
其富含于人心脏、肝脏、肾脏和胰腺中。人体内总含量为0.5~1.5g。在细胞内的分布为:细胞核内占25%~30%, 线粒体内占40%~50%, 微粒体内占15%~20%, 细胞质内占5%~10%。
辅酶Q10作为人体最重要的辅酶之一, 辅酶Q10的主要功能有:自由基清除剂、抗肿瘤药物、增强免疫力(机体非特异性免疫增强剂) 、代谢激活剂, 在细胞能量转换中具重要作用、抗氧化作用、增强心脏对缺氧的忍耐力等。目前, 辅酶Q10作为多功能生化药物在临床上、作为添加剂在食品中和化妆品中已有很多应用。随着对其应用研究的不断深入, 其应用范围将进一步扩大, 开发生产具有广阔的应用前景。
辅酶Q10 是体内组成呼吸链的必需成分 , 是电子传递链中的递氢体 , 为线粒体合成能量物质ATP的必要成分 , 主要来源于食物补充和自身合成。近年来越来越多的研究证明 , 还原性辅酶Q10 可以通过与维生素E协同作用清除自由基而在体内起抗氧化作用 , 是体内一种重要的脂溶性抗氧化剂。辅酶Q10 通过阻止体内LDL氧化修饰而对动脉粥样硬化的发生和发展有一定的抑制作用 , 但其作用机制尚不确切。
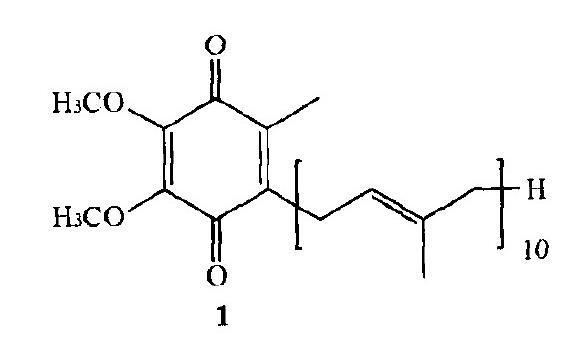
辅酶Q10,化学名称为2, 3-二甲氧基-5-甲基-6-癸异戊烯基苯醌,是真核细胞线粒体中电子传递链和有氧呼吸的参与物质之一,为黄色至橙黄色结晶性粉末,无臭无味,遇光易分解。
性状状为黄色至橙黄色结晶性粉末,无臭无味,遇光易分解。在三氯甲烷或丙酮中溶解,在乙醇中极微溶解,在水中不溶。熔点为48~52°C。
主要制剂有四种:(1)辅酶Q10片;(2)辅酶Q10软胶囊;(3)辅酶Q10注射液;(4)辅酶Q10胶囊。
一、辅酶Q10的发现
1957年, 英国Morton教授从患有维生素A缺乏症的大鼠肝脏中分离得到辅酶Q10(辅酶Q10) , 并称之为辅酶q10, 意即广泛存在的醌。1958年, Folkers等不仅阐明了辅酶Q10的化学结构, 还进行了辅酶Q10的化学全合成, 并首次通过发酵手段获得了辅酶Q10。1972年, 意大利Littarru等发表了心脏病患者缺乏辅酶Q10的研究报告。到了70年代中期, Mitchell用化学渗透假说理论, 揭示了生物体内能量的转换以及辅酶Q10在能量转换体系中重要的质子传递作用。80年代初, 随着瑞典Ernster揭示出辅酶Q10的抗氧化作用和自由基清除作用, 特别是日本实现了批量生产纯品辅酶Q10的工业技术, 极大地满足了大量临床试验对辅酶Q10的需求, 有力地促进了辅酶Q10临床研究的进展。同时由于色谱分析手段的应用, 实现了用高效液相色谱仪直接检测血液和组织中的辅酶Q10含量, 使得辅酶Q10的临床试验在数量和规模上都得到了迅猛发展。
二、辅酶Q10的现代研究进展
2.1 药理
辅酶Q10具有促进氧化磷酸化反应和保护生物膜结构完整性的功能。辅酶Q是生物体内广泛存在的脂溶性醌类化合物, 不同来源的辅酶Q其侧链异戊烯单位的数目不同, 人类和哺乳动物是10个异戊烯单位, 故称辅酶Q10。辅酶Q在体内呼吸链中质子移位及电子传递中起重要作用, 它是细胞呼吸和细胞代谢的激活剂, 也是重要的抗氧化剂和非特异性免疫增强剂。
2.1.1 自由基清除作用
过量自由基严重影响左心室功能, 超氧化物歧化酶(SOD) 可明显改善心室功能。Nayler在动物实验中发现辅酶Q1 0与S O D有类似作用。采用Langendorff方法研究了辅酶Q10对缺血性心肌的保护作用, 发现辅酶Q10可明显降低线粒体氧耗, 减少细胞三磷酸腺苷消耗。超微结构检查证实, 辅酶Q10能维持细胞线粒体结构的完整性, 防止细胞水肿, 细胞膜破裂, 以及线粒体溶解与肌纤维的紊乱性排列。Ohhara与Sugiyaman分别在大白鼠及狗中证实了辅酶Q10的这些作用, 并认为辅酶Q10可提高心肌张力恢复的百分率。
近几年来许多实验均证实辅酶Q10可明显降低心肌缺血再灌注后丙二醛(M D A) 的生成, 减轻线粒体肿胀, 保持细胞膜结构的完整性, 缩小心肌缺血范围。这些均表明辅酶Q10具有抗氧自由基, 维持细胞结构的完整性从而达到保护心肌的作用。
孙晓芳等研究了辅酶Q10抗衰老小鼠的脂质过氧化作用。其采用D-半乳糖制备衰老小鼠模型, 每日灌胃给予辅酶Q10。试验结果表明, 辅酶Q10可使肾、肝、睾丸组织超氧化物岐化酶的活性升高, 过氧化脂质的分解产物含量下降。因此证实, 辅酶Q10可增强这些器官对自由基的清除能力, 抑制脂质过氧化, 减少其分解产物含量, 起到保护这些器官的作用, 从而可以对抗D-半乳糖所致衰老退行性病变。
王怀颖等也采用D-半乳糖衰老模型小鼠, 研究了辅酶Q10对老化模型小鼠大脑皮层抗氧化功能的影响, 结果表明辅酶Q10能提高脑组织的抗氧化能力, 其作用与维生素E无显著性差异。
2.1.2 稳定细胞膜维持钙离子通道完整
Ichihara指出, 心肌缺血、缺氧时, 由于心肌细胞膜受到损伤导致膜内pH下降, 并失去了结合Ca2+能力, 大量Ca2+以丛状形式沉积于线粒体, 即造成了所谓的Ca2+负荷过重。Kobayashi认为, 辅酶Q10能增加ATP合成, 具有直接保护细胞膜稳定细胞内外环境的作用。有的学者认为, 辅酶Q10对缺血时心肌细胞线粒体畸变的防止作用, 也是辅酶Q10维持钙离子通道完整的佐证。
2.1.3 纠正病变组织辅酶Q10的缺乏状态
Folkers发现几乎所有心脏病患者以及某些非心血管疾病患者, 其组织中均有内源性辅酶Q1 0缺乏, 而给予外源性辅酶Q10可使患者临床症状明显改善。此外, Folker在对1000例各种病因所致的心脏病患者的测试中发现, 这些患者体内辅酶Q10水平明显低于正常人。
Littaru等报告132例心脏病患者中有98例辅酶Q10含量较正常水平低25%以上, 三尖瓣缺损者辅酶Q10缺乏最严重, 较正常水平低82%。Ⅰ或Ⅱ级心脏病患者辅酶Q10缺乏较少, Ⅲ或Ⅳ级心脏病患者辅酶Q10缺乏最为明显。Watanabe等报告了406例心脏病患者的血液研究结果也表明许多患者缺乏辅酶Q10
2.1.4 免疫增强作用
Bliznakov等曾以细菌或白血病病毒感染小鼠, 发现辅酶Q10可增强小鼠的免疫反应及白细胞的吞噬能力, 提高宿主对感染的防御机能, 并可延长其存活时间。
2.1.5 辅酶Q10促进动物学习记忆
姚文兵等利用东莨菪碱所致记忆障碍模型小鼠和N-甲基-D-天冬氨酸(NMDA) 单侧损伤Meynert基底核大鼠, 采用放射化学法测定大鼠皮层乙酰胆碱转移酶(ChAT) 活性, 以观察辅酶Q1 0对东莨菪碱所致记忆障碍模型小鼠和NMDA单侧损伤Meynert基底核大鼠学习记忆的促进作用。结果辅酶Q10对东莨菪碱所致学习记忆损害具有显著的改善作用, 能显著的提高损伤大鼠空间辨别学习记忆能力;对东莨菪碱所致的小鼠脑内胆碱酯酶活性增高无拮抗作用, 但能显著增高损伤小鼠皮层ChAT活性。表明辅酶Q10对动物学习记忆有明显促进作用。
张国平等观察了微循环障碍的大鼠, 在全身缺血缺氧的情况下, 经辅酶Q10 3 d灌服后肠系膜微循环的变化。结果发现, 在血压变化不显著的同时, 辅酶Q10对微血管口径无明显影响, 但对血流有突出的改善作用, 其表现为, 实验组大鼠毛细血管中红细胞流速明显加快, 红细胞聚集明显减轻, 白细胞贴壁黏附显著减少, 这些变化均可使微循环血流更加通畅, 循环加快, 使氧的转移、释放增加, 因而改善了微循环障碍大鼠体内的缺氧状态。结果表明辅酶Q10具有治疗微循环障碍, 改善细胞缺氧, 减轻细胞损伤的作用。
2.1.6 药动学研究
口服辅酶Q10 100 mg后, 血浆药物浓度在5~10 h(平均6.5 h) 达峰值, 血浆平均浓度为(1.004±0.3) mg/L;一日3次, 每次服用100 mg后的平均稳态血药浓度为5.4 mg/L, 血浆半衰期为(33.9±5.32) h, 显示该药代谢及消除均缓慢。辅酶Q10从胃肠道吸收后被乳糜微粒摄取, 大部分积存于肝内并转化为极低密度脂蛋白(V L D L) , 此后在体内某些器官如肾上腺、肝、肺、肾和心肌组织中分布。多数患者用药后24 h血浆浓度出现两次峰值, 推测为肝脏摄取的辅酶Q10向外周血循环释放再分布所致。辅酶Q10从胆道排泄, 由粪便排出体外。
2.2 毒理
本品急性毒性实验, 小白鼠及大白鼠LD50均大于4 000 mg/kg。在亚急性毒性实验中, 给予Wistar大白鼠每日40, 200及1 000 mg/kg的剂量, 共用5周;雌雄家兔每日6, 60及600 mg/kg, 连续应用23 d, 受试动物的血液及尿液检查及形态观察均无异常变化。在慢性毒性实验中, 给予Wistar大白鼠每日6, 60及600 mg/kg的剂量, 连续26周, 受试动物的一般状态、血液和尿检查、形态观察均无异常改变。
2.3 应用
2.3.1 充血性心力衰竭
1970年Folkers首次发现充血性心力衰竭患者心肌内的辅酶Q10的含量降低, 于是提出了辅酶Q10治疗充血性心力衰竭的基本理论。后来他通过1 000余例的心肌活检证实充血性心力衰竭患者内源性辅酶Q10含量明显低于正常人, 而且发现心衰程度愈重, 心肌内辅酶Q10含量愈低, 辅酶Q10的治疗效果愈好。Langsjoen等对19名Ⅲ~Ⅳ期充血性心衰病人辅酶Q10治疗, 结果表明在治疗期间, 平均射血分数和心搏量增加。
2.3.2 心律失常
Hosono等报告, 辅酶Q10使培养的离体心肌细胞有规则节律比例增加, 节律不齐发生频率减少。对冠心病、风湿性心肌炎、病毒性心肌炎等所致房性早搏、室性早搏、阵发性房颤均有效。国内以心律不齐为代表性病种, 采用双盲法验证辅酶Q10对心脏疾病的疗效, 按照1979年全国冠心病中西医结合治疗座谈会制定的诊断标准, 选择106名例室性早搏患者, 给药组5 6例, 口服辅酶Q1 0, 每次20 mg, 一日3次。对照组50例, 口服安慰剂胶囊。治疗40 d后给药组总有效率达87.5%, 67%的患者心电图恢复正常, 8 5%心悸、胸闷、头昏等症状消失;而对照组的有效率为22%, 绝大多数患者的心电图、临床症状没有改善, 经统计学处理两组有显著性差异(P<0.01)。
2.3.3 心绞痛
β-受体阻滞剂、有机硝酸酯类及钙通道阻滞剂为迄今为止公认有效的治疗心绞痛的药物。近年的动物试验和临床研究资料表明, 辅酶Q10对稳定型心绞痛有相当的疗效, 而对变异型心绞痛的疗效尚未见报道。Kamikawa通过随机双盲交叉临床验证, 稳定型心绞痛患者口服辅酶Q1030 mg/d, 疗程1~8周, 大部分患者感到胸部压迫感, 气短和心悸等症状得到改善, 心绞痛发作次数减少, 含服硝基甘油量较对照组少, 运动耐量显著提高。最近, 美国采用辅酶Q10治疗冠心病心绞痛进入广泛临床研究, 分别使用150 mg/d和300 mg/d两种剂量的辅酶Q10进行治疗, 并与安慰剂对照, 结果证实辅酶Q10治疗组缓解心绞痛和提高运动耐量均明显优于安慰剂组, 而且临床疗效与药物剂量呈线性关系。
2.3.4 高血压
Yamagami等发现原发性高血压患者白细胞内的辅酶Q10酶活性降低, 于是对20例血清辅酶Q10降低的高血压病人服用辅酶Q10 33 mg, 1日3次, 共12周, 结果收缩压和舒张压均有明显降低。有人报道, 原发性高血压患者的血淋巴细胞中显著缺乏辅酶Q1 0, 予以外源性辅酶Q1 0后可纠正, 但停药后又出现缺乏。山上彻统计72例用辅酶Q10治疗的高血压患者, 其收缩压由(180.1 1±23.20) mmHg降到(165.68±23.28) mmHg(P<0.01) , 舒张压由(98.83±15.63) mmHg降到(91.89±14.09) mmHg(P<0.01) 。国内报导的辅酶Q10治疗高血压的临床效果不一, 最低有效率为18%, 最高为100%。辅酶Q10主要适用于缺乏辅酶Q10所致的原发性高血压。目前临床上常用的8种降压药物, 其中除利血平外, 均会增加高血压患者心脏中辅酶Q10的缺乏。使用降压药物时并用辅酶Q10, 可增强降压效果, 预防不良反应发生。
2.3.5 慢性肝炎
辅酶Q10可作为免疫功能调整药而用于治疗慢性肝炎。北京医学院附属第一医院治疗慢性病毒性肝炎50例, 血清谷丙转氨酶平均下降19.7%, 而对照组却上升3.28%, 治疗组显效率(32%) 明显高于对照组显效率(10%) 。江苏省人民医院等8家医疗单位验证209例, 治疗慢性肝炎显效率75%, 有效率9 0%。
2.3.6 抗肿瘤
通过测定癌症患者体内辅酶Q10浓度发现, 与正常人相比其含量较低, 1994年Lockwood等研究了各种各样癌症患者包括乳房癌、前列腺癌、胰癌和结肠癌等患者, 对其体内辅酶Q10缺乏发生率进行研究, 对32位已转移至腋窝淋巴结的乳房癌患者, 经服用辅酶Q10 90 mg/d并结合服用一定剂量的抗氧化剂如维生素C、维生素E、ß-胡萝卜素等不饱和脂肪酸, 经1年试验, 患者均未发生肿瘤转移, 其中6例其肿瘤发生部分消退。
2.3.7 增强免疫力
1970年, 首先报道了老鼠服用了辅酶Q10能提高机体免疫细胞杀死细菌的活力, 并提高抗体反应;辅酶Q1 0能增加白细胞的数量, 增加胸腺活力, 激发免疫球蛋白和抗体数量增加。1 9 8 5年Folkers等研究了获得性免疫缺陷症(AIDS) 患者, 其血清中显示出较低的辅酶Q10浓度, 同时在临床上对8位成年患者经60 mg/d辅酶Q10治疗, 一个月后, 血清IgG含量明显提高。因此辅酶Q10作为机体非特异性免疫增强剂在肿瘤提高机体免疫力等方面可作为一种较好的治疗药物。
2.3.8 治疗帕金森病
美国加州大学San Diego分校医学院的Clifford Shults指出, 根据一项多中心、双盲试验的结果, 辅酶Q10能减慢帕金森病的发展。
赵春玉等用辅酶Q10治疗20例帕金森病患者, 取得了可喜的效果。治疗组用辅酶Q10和多巴丝肼治疗6个月后停服辅酶Q10, 再只用多巴丝肼治疗6个月。停用辅酶Q10后出现病情加重或必须加大多巴丝肼用量者视为有效病例。治疗组有效率为66.7%。
4 不良反应
临床剂量的辅酶Q10迄今为止尚未发现严重的不良反应。主要不良反应有上腹不适、恶心、食欲不振和腹泻, 偶有荨麻疹和一过性心悸。辅酶Q10与其它药物一般不发生作用, 某些降糖药可能干扰辅酶Q10的吸收和利用, 但需进一步观察。有人曾作过辅酶Q10急性、亚急性和慢性毒性试验, 对被检查的一般症状、血液学、及血液生化等14项指标, 解剖观察以心、肝、肺、脾、肾等器官的组织学检查, 均未发现辅酶Q10引起的病理变化和毒性反应。在国内外多年的临床研究报告中, 尚未发现因使用辅酶Q1 0而出现明显毒副反应的病例。辅酶Q1 0不是异体物质, 对人体是无害的。
三、辅酶Q10缺乏综合征
辅酶Q10缺乏综合征是以儿童时期发病为主的、罕见的、多系统受累的一组临床综合征,主要表现为:脑肌病、严重的婴儿多系统疾病、肾脏病变、小脑性共济失调和孤立性肌病。本文通过对近年来文献的回顾,针对辅酶Q10缺乏综合征的分类、发病机制、临床表现、治疗和展望作一综述。
辅酶Q10缺乏综合征(Coenzyme Q10Deficiency Syndrome)是一种罕见的临床和遗传异质性综合征,迄今报道约100余例。目前认为它有5种主要的临床表型:脑肌病、婴儿严重的多系统疾病、肾脏病变、小脑性共济失调和孤立性肌病。其中小脑性共济失调和综合征性或孤立性肾病综合征是最常见的形式。虽然小儿患者比较常见,但实际上可在任何年龄发病。迄今为止,仅在很少部分病人中发现致病突变,因此很难确定相关表型与基因型之间的关联。因为辅酶Q10缺乏综合征(特别是肌肉症状和肾病)患者通常对补充辅酶Q10反应良好,所以该病的早期识别极为重要。
3.1 分类
3.2.1 AHM的作用
本病根据突变基因是否直接参与辅酶Q10生物合成,可以分为原发性辅酶Q10缺乏综合征和继发性辅酶Q10缺乏综合征。
3.1.1 原发性辅酶Q10缺乏综合征
研究发现,人类至少有13个基因与辅酶Q10的生物合成相关,这些基因突变被认为与原发性辅酶Q10缺乏综合征的发病相关。迄今,这些基因中的8个突变(PDSS1、PDSS2、COQ2、COQ4、COQ6、ADCK3、ADCK4和COQ9)已被报道,这些突变可引起各种临床表型,从致命的婴儿多系统疾病到成人的脑病。
3.1.2 继发性辅酶Q10缺乏综合征
辅酶Q10缺乏综合征也可作为继发表现,在其他疾病中见到。通过对与辅酶Q10生物合成无关的基因突变患者进行肌肉或皮肤成纤维细胞的检测,发现在肌肉和成纤维细胞内辅酶Q10水平下降。这些突变基因包括:APTX基因、BRAF基因、ETFDH基因和许多mtDNA的突变。Haas等人已在甲基丙二酸尿症的患者细胞中观察到辅酶Q10缺乏,但二者的内在关联并不清楚。
3.2 发病机制
辅酶Q10缺乏综合征的发病机制仍不清楚。目前认为它可能与生物能量代谢缺陷相关,但也可能与除直接参与线粒体ATP合成外的、辅酶Q10的其他生理功能异常相关,如嘧啶代谢受损、活性氧增加和自噬异常等。目前多数学者认为辅酶Q10缺乏,导致线粒体呼吸链的复合物之间电子传递障碍,这使得本病患者临床表现类似于其他呼吸链疾病(脑肌病、共济失调、乳酸性酸中毒、感觉神经性耳聋、视网膜色素变性和肥厚性心肌病)。但部分表现则是辅酶Q10缺乏综合征特有的,如激素抵抗型肾病综合征(SRNS),一般不发生在呼吸链疾病中。
3.3 临床表现
该临床表型的主要表现是线粒体肌病、复发性肌红蛋白尿和脑病的临床三联征,肌肉活检发现显著降低的辅酶Q10浓度,可见于ADCK3基因突变。
Ogasahara等人于1989年首次提出辅酶Q10缺乏症。他们发现:同一家系中的2姐妹早期发育正常,临床表现为运动不耐受和缓慢进展的中轴和四肢近端肌肉无力。患者在5岁时出现脑部受累表现:姐妹均出现学习障碍,其中1例出现癫痫发作,而另1例出现小脑相关症状。此外,2例均在癫痫发作或并发感染后出现肌红蛋白尿。实验室检查异常包括乳酸性酸中毒和血清肌酸激酶升高;肌电图显示肌病的特点。肌肉活检显示Ⅰ型肌纤维内出现破碎红纤维和过多的脂滴堆积,同时发现降低的线粒体呼吸链复合物Ⅰ+Ⅲ和Ⅱ+Ⅲ的活性,以及显著降低的辅酶Q10浓度(约为正常人的5%)。与此相反,在患者血清和培养的成纤维细胞中的辅酶Q10水平正常。
此后,又有几例患者出现相似的线粒体肌病、复发性肌红蛋白尿和脑病的临床三联征,且在其中1例患者基因检测时发现ADCK3基因突变。在补充辅酶Q10治疗后,患者肌肉症状得到不同程度的改善。
3.3.2 严重的婴儿多系统疾病
该临床表型有广泛的表现,可累及肝、肾、心脏和脑等器官,出现呼吸功能不全、肾病综合征、乳酸性酸中毒、肥厚型心肌病、听力丧失、癫痫发作和糖尿病等表现,可见于COQ2、COQ9、PDSS1和PDSS2基因突变。
3.3.2.1 COQ2基因突变
Rtig等人于2000年在2例同胞的基因检测中发现在COQ2基因的外显子7区出现纯合碱基对的缺失。其中妹妹出现新生儿神经性窘迫、肾病综合征、肝病、全血细胞减少、糖尿病、癫痫发作和乳酸性酸中毒,在第12天时进展为致死性多器官衰竭。哥哥也出现贫血、肝功能衰竭和肾功能不全,在出生1d后死亡。
Jakobs等人在2013年报道2例纯合COQ2突变,来自土耳其近亲结婚的父母生育的异卵双胞胎均患病。患儿出现早产、呼吸功能不全、癫痫发作、喂养困难、发作性呼吸暂停、全身性肌张力下降和全身性水肿,分别在5~6个月龄时死亡。与Dinwiddie等人的发现一样,这对双胞胎并无肾脏受累的证据。
最近几年,在其他数例出现重度婴儿多系统病变的患儿的基因检测中同样发现COQ2基因突变。此外,成纤维细胞内的辅酶Q10缺乏和早期肾脏受累似乎是原发性婴儿多系统综合征的一个标志。
3.3.2.2 COQ9基因突变
Rahman等人在2001年报道1例新生儿出现广泛的四肢肌张力增高、躯干肌肉张力下降、乳酸性酸中毒、肾小管病变、左心室肥厚及全身性运动减少。头部MRI检查显示大脑和小脑萎缩。该患儿随后出现严重的癫痫发作和肌张力障碍,并在2岁时死亡。通过纯合子定位和候选基因筛查发现COQ9纯合突变。
3.3.2.3 PDSS1基因突变
Mollet等人2007年在1个近亲结婚的家系中,发现2兄妹均表现为辅酶Q10缺乏、早发性耳聋、脑病、肥胖、网状青斑和心脏瓣膜病。基因检测发现纯合PDSS1基因突变。
3.3.2.4 PDSS2基因突变
Rtig等人2000年时对1个家系中的姐弟3人研究发现:患者在出生后迅速出现神经系统症状,包括:眼球震颤、视神经萎缩、感觉性神经性听力丧失、共济失调、肌张力障碍、乏力、急进性肾病和广泛的辅酶Q10缺乏。进行基因检测后发现PDSS2基因突变。López等人在2006年报道1例出现肾病综合征和Leigh综合征的男婴。该患儿表现为新生儿肺炎和肌张力低下。在3个月时出现癫痫发作、进展性软瘫,伴喂养困难、严重的阵发性呕吐和乳酸性酸中毒,在8个月时死于癫痫持续状态。该患儿基因检测发现PDSS2基因突变,该基因编码聚异戊二烯焦磷酸合酶的2个子单元中的1个,该酶是辅酶Q10的生物合成途径的限速酶。
3.3.3 肾脏病变
该临床表型的主要表现是伴或不伴感觉性听力丧失的孤立性肾病,部分患者表现为激素抵抗型肾病综合征,可见于COQ2和ADCK4基因突变。
Salviati等人2005年首次发现在携带COQ2基因突变的2姐弟中,姐姐在12个月时出现肾病综合征,不伴神经病学改变。在2007年,Diomedi-Camassei等人描述另外2例COQ2基因突变患儿出现早发性肾小球疾病。其中第1例患儿在18个月龄时,由于塌陷性肾小球疾病出现激素抵抗型肾病综合征,不伴肾外表现。第2例患儿在第5天时出现少尿表现,肾穿刺活检发现严重的毛细血管外增生,迅速发展为终末期肾病,最终出现进展性癫痫性脑病,在第6月龄时死亡。同年,Diomedi-Camassei等人在该患儿的肾皮质和骨骼肌的检测发现呼吸链复合物Ⅱ+Ⅲ活性和辅酶Q10水平的降低。
McCarthy等人在2013年采用新一代测序技术对36例激素抵抗型肾病综合征患儿进行相关的24个基因同时筛选,结果发现其中1例患儿的COQ2基因发生2种突变。该患儿在2岁时出现肾脏疾病和急性肾功能不全,不伴神经系统症状和感觉性神经性听力下降。另外,Mitsui等人在新英格兰杂志上曾报道同1家系中2例COQ2基因突变患者表现为迟发性激素抵抗型肾病综合征。这2例病人在60~70岁时出现多系统萎缩、色素性视网膜炎和肾脏病变。因此,有学者认为COQ2基因的多态性可能与多系统萎缩相关,但该假设尚需验证。
同年,Ashraf等人在7个家系中的14例病人中发现ADCK4基因突变,主要表现为激素抵抗型肾病综合征。除1例病人在1岁时发病、表现为发育迟缓外,患者多在十几岁时发病。其中仅1例病人对补充辅酶Q10治疗反应良好。
3.3.4 小脑性共济失调
辅酶Q10缺乏综合征的小脑型最常见,特点是小脑性共济失调和小脑萎缩,且与神经病变、癫痫、智力低下、肌肉无力、性腺功能减退和成纤维细胞内辅酶Q10水平降低广泛相关。肌肉形态学检查未发现破碎红纤维和脂质沉积性肌病。部分患儿携带APTX或ADCK3/CABC1基因突变。
据Horvath等在2012年报道:除2例成年起病的、表现为小脑性共济失调和性腺功能减退的患者及2例成年起病的、伴ADCK3基因突变的、临床表型较轻的患者外,其余患者均为儿童期或青少年期起病。有趣的是,在肌肉未严重受累的患者,辅酶Q10水平是正常的。在其他出现小脑性共济失调、小脑萎缩、运动不耐受、肌张力障碍和轻度认知障碍的患者发现了ADCK3基因突变。但并未进一步研究这些患者肌肉和成纤维细胞中的辅酶Q10水平。
3.3.5 孤立性肌病
该临床表型主要表现为单纯的肌病,不伴其他系统受累表现,肌肉病理发现脂质沉积和线粒体肌病的表现,目前认为与ETFDH基因突变有关。
Lalani等人和Horvath等人分别在2005年和2006年分别报道了1例辅酶Q10缺乏综合征患者仅有肌病的表型,进一步检测发现肌肉内脂质沉积和呼吸链功能障碍。
Gempel等人在2007年对5个家系的7例出现运动不耐受、疲劳、近端肌病和血清肌酸激酶升高的患者进行基因检测,发现编码电子传递黄素蛋白脱氢酶(先前认为与Ⅱ型戊二酸尿症相关)的ETFDH基因突变。对肌肉进行免疫组织化学染色发现脂质沉积和线粒体肌病的表现。其中单纯性肌病的患者在补充辅酶Q10后均出现显著缓解。相比之下,其他研究报道的出现多种酰基辅酶A脱氢酶缺乏症和ETFDH基因突变患者肌肉检测发现辅酶Q10水平正常。
3.4 治疗与展望
目前关于本病的治疗尚缺乏统一的认识。但早期进行肌肉的复合物Ⅰ+Ⅲ和Ⅱ+Ⅲ活性和辅酶Q10水平的检测,及时补充辅酶Q10治疗是改善患者预后的关键。多数研究者认为:早期治疗可在一定程度上缓解患者病情和阻止各系统症状和体征的发展。如不及时治疗,一旦造成关键器官(如肾脏、脑等)的损伤,难以通过治疗得到修复。另外,患者的生存期差别很大,从出生后数天死亡到老年期死亡的病例均有报道,这可能与本病的临床和遗传异质性相关。随着对本病发病机制的探索、基因型与表型之间相关性的研究和补充辅酶Q10及其类似物疗效评价,必定会为辅酶Q10缺乏综合征患者提供更有效的治疗方案。
四、辅酶Q10对卵巢储备功能下降患者的防治作用
卵巢储备功能下降病理上主要表现为卵巢皮质内原始卵泡数量过早耗竭,卵母细胞质量进行性下降,临床上表现为激素分泌异常及生育力的持续下降等。线粒体与卵巢储备功能密切关联。辅酶Q10参与线粒体呼吸链中的电子转运和氧化磷酸化,具有清除自由基和抑制细胞凋亡的作用,辅酶Q10可以通过改善线粒体功能,对抗线粒体卵巢衰老和生理程序性卵巢衰老来保护卵巢储备。辅酶Q10预治疗可以增强卵巢对刺激的反应,改善卵巢储备下降患者的卵母细胞和胚胎质量。对于卵巢储备功能下降患者给予一段时间的辅酶Q10预治疗对其卵巢功能恢复和未来妊娠有明显效果。
辅酶Q10于1957年被发现。1958年,美国德克萨斯大学的Karufulukas博士发现了它的化学结构。人体内的辅酶Q10含量随着年龄的增长而降低,辅酶Q10水平降低会影响线粒体的氧化磷酸化,减少细胞能量物质三磷酸腺苷(adenosine triphosphate,ATP)的生成,影响卵子质量和卵子受精。近年来,随着二孩政策的实施,女性的备孕年龄上升,以及环境污染、社会压力等外在因素的影响,导致卵巢储备功能下降(decreased ovarian reserve,DOR)的患者逐渐增多。辅酶Q10作为线粒体内膜内的质子和电子载体,具有生物能量作用,同时也是一种内源性亲脂性抗氧化剂,广泛存在于生物膜中,主要参与线粒体呼吸链中的电子转运和氧化磷酸化,可以抗氧化、清除自由基,提高卵巢储备功能,增加卵泡数量,改善卵泡质量。笔者概述辅酶Q10对DOR患者的防治作用及其机制。
4.1 DOR
美国疾病预防控制中心辅助生育协会(SART-CDC)提出:由于年龄、遗传因素、医源性或手术原因导致的卵巢内存留卵子的质量和数量下降,被定义为DOR。DOR的检测指标:卵泡刺激素(follicle stimulating homone,FSH)由垂体前叶的促性腺激素分泌,促进卵泡发育。血清FSH水平随年龄增加而增加,早卵泡期血清FSH水平>10 U·L-1是DOR的预测指标。此外,抗苗勒激素(anti-mulerian hormone,AMH)、窦卵泡计数(antral follicle count,AFC)、抑制素B(inhibin B,INHB)、卵巢体积也用于预测卵巢储备。AMH是卵巢储备的重要标志,在卵泡发育和卵泡选择中起着重要作用。一般认为血清基础AMH水平0.5~1.0 ng·mL-1,预示着DOR。INHB是小窦卵泡生长的标志物,可在血清基础FSH第3天升高前预测DOR。
DOR病理上主要表现为卵巢皮质内原始卵泡数量过早耗竭,卵母细胞质量进行性下降,临床上表现为激素分泌异常及生育力的持续下降,如雌激素降低、AMH降低、FSH升高等。关于DOR病因尚无统一定论,主要包括年龄、遗传因素(某些性染色体缺陷和常染色体基因缺陷)、代谢免疫因素、心理环境因素及医源性因素等。近几十年来,辅助生殖技术飞速发展,为治疗不孕、不育提供了非常有效的方法,但DOR患者在接受辅助生殖技术助孕过程中,常常出现卵巢低反应、促排卵药物用量增加、高周期取消率、低临床妊娠率、低抱婴率等情况。因此,如何提高DOR患者辅助生殖妊娠结局成为困扰临床医疗工作的难题。
4.2 线粒体对卵巢储备功能的影响
线粒体是存在于大多数细胞中的细胞器,被两层膜包被,是细胞进行氧化磷酸化和产生能量的主要场所。线粒体根据不同的生物种类及生理状态而在细胞中呈现不同形状。线粒体主要影响细胞的能量转化,其次亦参与其他的生理功能:如调控细胞增殖与代谢,调节细胞凋亡过程,调节钙离子浓度,合成胆固醇及某些血红素等。DOR除了表现为卵泡数量减少外,主要还表现为卵子质量下降,而卵子质量与线粒体功能密切相关,线粒体阻止卵子的凋亡,促进胚胎的形成,其数量影响ATP的生成。线粒体产生的ATP是触发卵子受精必不可少的条件。有文献报道,在卵母细胞、胚胎形成及发育过程中,线粒体参与钙离子的储存与转运,线粒体损伤导致卵子异常受精;线粒体功能异常会驱动凋亡途径发生,细胞过度氧化应激(oxidative stress,OS),影响优质胚胎的形成。
线粒体为卵母细胞成熟、受精和卵裂提供能量。早期卵泡衰竭及细胞凋亡是DOR发生的机制。线粒体基因异常(数量变化、mtDNA片段缺失、突变)会降低细胞内的能量生成。有学者认为,低ATP水平可以激活卵子中哺乳动物雷帕霉素靶蛋白(mTOR)信号通路,导致原始卵泡的异常活化、丢失增多,卵泡过早衰竭,逐渐发展为DOR。线粒体参与细胞的各项生物代谢过程,研究表明,线粒体可以作为评估卵母细胞发育潜能的指标,线粒体异常是导致高龄不孕患者卵母细胞发育潜能低下的关键因素。
人类的所有细胞中都含有线粒体,而卵母细胞被认为是最需要线粒体的细胞,mtDNA拷贝数是评估卵母细胞活性的重要参考指标。在颗粒细胞、卵母细胞或外周血均能发现mtDNA拷贝数与卵巢储备功能的关系。TSAI等发现卵子颗粒细胞mtDNA拷贝数影响辅助生殖治疗周期的结果。研究表明,线粒体数量异常可导致卵子胞浆成熟障碍及卵细胞受精异常。卵细胞及胚胎发育过程中,线粒体存在一个“阈值”,当线粒体数量高于阈值时,卵细胞和胚胎发育良好。线粒体数量的减少,直接导致能量合成障碍,影响颗粒细胞功能。辅助生殖促排卵过程中退化的卵母细胞中线粒体数量较少。与正常卵巢储备比较,DOR组卵母细胞线粒体数量明显减少。对接受胚胎植入前基因诊断患者的极体和卵裂球进行分析,发现高龄女性卵母细胞极体含mtDNA拷贝数少于年轻女性。国内学者采用实时荧光定量聚合酶链反应(PCR)技术检测不同受精结局的卵母细胞,证实受精能力与mtDNA拷贝数有关。DIOGUARDI等在卵巢储备功能下降的小鼠中,也发现颗粒细胞及卵母细胞mtDNA拷贝数降低。此外,有研究表明,对卵巢储备功能不同的患者进行外周血mtDNA拷贝数检测,发现DOR患者mtDNA含量低于卵巢储备正常者。
线粒体阈值数量可以保障卵母细胞成熟及功能,线粒体数量不足导致卵母细胞发育和成熟异常,这可能是DOR患者卵泡数量减少及质量下降的病因之一。
卵巢储备功能表现了女性的生育能力,卵子质量的优劣决定了胚胎的质量。随着空气污染及盆腔附件侵入性操作增多等,由于DOR而不孕的女性数量逐年上升。目前对DOR病因探索研究仍是一个热点。氧化磷酸化离不开线粒体,由于线粒体缺乏保护及严格的修复机制,线粒体数量不足、mtDNA片段缺失、突变都可以引起功能发生变化,最终导致能量产生减少。
4.3 辅酶Q10、线粒体与卵巢储备功能
辅酶Q10具有清除自由基、抗凋亡、稳定细胞膜、减少OS、提高机体免疫力等优点,是重要的抗氧化剂和免疫增强剂,是线粒体电子传递链的重要组成部分,起到能量转换剂和明显的抗脂质过氧化作用。在临床上,它具有抗衰老、抗疲劳、抗高血压和保护心脏等作用,具有无毒、无致畸和无明显不良反应的优点,越来越多受到基础研究者和临床工作者的关注。
辅酶Q10主要参与线粒体呼吸链中的电子转运和氧化磷酸化,产生ATP,通过抑制脂质过氧化和DNA氧化作用,增强细胞内的内源性抗氧化系统。补充辅酶Q10已被证明可以改善心血管功能和男性生育能力。血浆中辅酶Q10浓度降低与性腺机能减退和其他类固醇激素水平改变有关。辅酶Q10水平的下降在35岁以上的人群中很常见,似乎与年龄相关的生育能力下降和胚胎非整倍体率的增加同时发生,这表明辅酶Q10表达的减少与卵巢老化有关。多项动物实验研究表明,辅酶Q10可以保护卵巢储备,通过恢复线粒体功能来抵消卵巢生理衰老,提高胚胎卵裂率和囊胚形成率。临床上,在35~ 43岁的妇女中,辅酶Q10预治疗可增强促排卵效果,降低胎儿非整倍性的概率。
辅酶Q10也通过调节细胞内部的氧化还原状态来影响基因表达。在生殖生物学中,辅酶Q10已被证明在男性和女性的生育能力中发挥作用。在特发性弱精子症不育症患者的精浆和精子细胞中,辅酶Q10水平降低和氧化增加。这些患者服用辅酶Q10可增加辅酶Q10含量,改善其氧化状态和精子运动特性。在雌性生殖系统中,外源性补充辅酶Q10的早期报告显示,在各种大鼠组织中,肾上腺和卵巢对辅酶Q10的摄取显著。这些器官的辅酶Q10总含量增加了1倍以上,而其他器官的辅酶Q10含量仅略有增加。STOIKOVIC等研究调查了辅酶Q10作为牛胚胎体外培养补充物质的作用,发现早期胚胎卵裂率、囊胚形成率、孵化率、囊胚扩张率显著提高,内质细胞体积增大。STOIKOVIC等还观察到辅酶Q10培养的胚胎中ATP含量增加。所有这些参数都表明胚胎质量有所提高。与此观察相符,BEN-MEIR等报道,辅酶Q10在生殖衰老过程中,可以恢复卵母细胞线粒体功能和生育能力,通过使用缺乏辅酶Q10合成的动物模型,强调了辅酶Q10在生殖衰老中的关键作用,并通过补充辅酶Q10可以挽救卵巢储备的减少。
辅酶Q10具有清除自由基作用。自由基是细胞正常的代谢产物,正常情况下,自由基的产生和清除处于动态平衡状态,一旦平衡被打破,体内自由基积聚可引起OS,导致DNA、蛋白质和脂质的过氧化,引起线粒体功能障碍和细胞凋亡。OS和线粒体功能障碍相互作用,形成恶性循环。OS可积聚大量自由基,破坏线粒体复合物Ⅰ,与此同时,线粒体复合物Ⅰ的缺陷会产生更多的自由基。辅酶Q10是人体内唯一天然可再生的脂溶性抗氧化剂。辅酶Q10通过氧化还原型结构的转换,可以清除自由基,减少OS,改善线粒体功能障碍。
辅酶Q10具有抑制细胞凋亡的作用。辅酶Q10可以保护线粒体 DNA,消除氧自由基,减少OS,防止过氧化损伤引起的线粒体DNA突变,维持线粒体呼吸链的完整性,减少细胞凋亡;辅酶Q10可以提高线粒体ATP生产效率,促进细胞生长和代谢,延缓细胞凋亡;辅酶Q10可以通过抑制线粒体渗透转化通路的打开来阻止线粒体去极化,抑制线粒体依赖性凋亡通路的激活,起到抗凋亡的作用;辅酶Q10可以抑制活性氧相关的凋亡通路,在肾近曲小管细胞中,辅酶Q10通过磷酸化p66shc机制,显著抑制尼古丁诱导的活性氧生成和细胞凋亡,进而激活抗氧化反应元件(antioxidant response element,ARE)及锰超氧化物歧化酶(manganese superoxide dismutase,MnSOD)的启动子,促使MnSOD清除活性氧,从而抑制凋亡。
辅酶Q10还可以改善线粒体功能,恢复线粒体的能量生产。卵母细胞的线粒体功能障碍导致氧化磷酸化减少和线粒体产生的ATP非最佳水平,这与生殖性能差密切相关,包括卵巢储备减少、卵母细胞质量差、受精异常和移植前胚胎发育紊乱。线粒体产生的能量对类固醇激素的生物合成、卵母细胞成熟、受精和早期胚胎发育都很重要。
4.4 辅酶Q10预治疗对卵巢功能的影响
成熟卵母细胞中辅酶Q10水平明显高于其在畸形卵母细胞中,其水平导致I~II级与III~IV级胚胎的比值明显提高。这一结论被AKARSU等证实,并可能突出辅酶Q10在保护卵泡脂蛋白免受氧化作用,保持其功能。此外,不同研究也指出补充辅酶Q10(600 mg·d-1),持续60 d,能改善卵巢储备下降女性的卵巢反应,在体外受精过程中,通过降低其对促性腺激素的需求,提高卵泡预刺激的反应能力。
ÖZCAN等认为,卵巢衰老和OS之间存在密切联系,而辅酶Q10可能有效地保护卵巢不受铂化物作用中的活性氧形成成分的影响。因此,虽然OS在女性不孕中的作用尚不清楚,但补充辅酶Q10可以通过改善线粒体功能,对抗线粒体卵巢衰老和生理程序性卵巢衰老来保护卵巢储备。
BURSTEIN等在衰老小鼠模型中比较了辅酶Q10、白藜芦醇和R-ALA蛋白在促排卵前18周内对排卵卵母细胞数量及卵母细胞线粒体功能的影响。结果表明,辅酶Q10预治疗显著增加排卵卵母细胞的数量,改善卵母细胞线粒体功能,并将老年小鼠体内的活性氧水平降低到与幼龄小鼠类似的水平。BEN-MEIR等研究了补充辅酶Q10、α硫辛酸和白藜芦醇对受损卵母细胞线粒体功能和卵巢储备的影响,这些线粒体生物能量的刺激物质被皮下注射到9个月大的小鼠体内,结果表明,补充辅酶Q10可保护卵巢储备,改善卵母细胞质量和排卵率,以及提高线粒体性能。此外,还能够使正常发育的配子比例增加。
XU等发现辅酶Q10预治疗后获得的卵母细胞数明显多于对照组。辅酶Q10组受精卵数和受精率数显著多于对照组,每例患者第3天优质胚胎数多于对照组。辅酶Q10组冷冻胚胎数和复苏移植胚胎周期数显著增加。在接受辅酶Q10治疗的女性中,每次新鲜胚胎移植周期的临床妊娠率和活产率分别为34.85%和31.82%,对照组分别为25.00%和21.88%。结果表明,辅酶Q10预治疗导致卵巢反应所需的促性腺激素总量显著减少,刺激持续时间缩短,雌激素峰值升高,卵母细胞数量增多。辅酶Q10预治疗可显著提高受精率和优质胚胎数。与对照组比较,辅酶Q10治疗组因未对刺激作出反应而取消的周期数明显较低,因胚胎发育失败而取消的周期数较少,冷冻胚胎的周期较多。
五、辅酶Q10与癌症治疗相关研究进展
辅酶Q10广泛存在于机体各组织中, 主要参与线粒体中ATP的生成及抗氧化还原作用。肿瘤和非肿瘤疾病中发现了辅酶Q10水平的变化, 而且辅酶Q10对许多涉及代谢的基因表达、细胞转运、转录调控、细胞信号转导都有影响, 使得辅酶Q10可能成为一个有效的基因调节靶点。许多临床研究发现癌症患者体内的辅酶Q10水平降低, 通过辅酶Q10的治疗可降低患者体内肿瘤标志物及炎症物质水平, 但目前仍需大量的随机对照试验去进一步证实辅酶Q10对于癌症的治疗效果、安全性及临床上最佳的治疗剂量。
癌症仍然是世界上主要的致死疾病之一, 许多癌症仍未发现较好的治疗方案。临床上目前多采取的放疗和化疗具有巨大的副作用, 许多癌症患者常常因为严重的副作用不能坚持治疗, 以及导致了患者的生活质量极大的下降。Co Q10(coenzyme Q10, Co Q10是一种生物醌化合物, 它参与了线粒体中呼吸链的氧化反应并增强了线粒体中ATP的合成, 以及可以转变成具有抗氧化活性的还原物(泛醇) 参与抗氧化作用。Co Q10的抗氧化作用可以减少受损细胞的自由基和活性氧的产生, 还可以通过氧化还原信号来控制细胞的能量代谢和死亡的调节。目前许多研究发现Co Q10对许多涉及代谢的基因表达、细胞转运、转录调控以及细胞信号转导都有影响。有研究发现在部分癌症患者体内的Co Q10水平有统计学差异, 因此有学者通过使用Co Q10来治疗癌症患者后发现其可以改善癌症患者的预后, 且其较小的副作用受到许多临床医生的关注。本文就目前临床上关于Co Q10用于癌症治疗的研究进行简要综述。
5.1 Co Q10的基础背景及相关临床应用
国家癌症中心发布的《2013年中国恶性肿瘤发病和死亡分析》文中提到, 2013年全国新发肿瘤病例数368.2万例, 肿瘤死亡数222.9万例。而且Chen等的研究数据显示2015年中国估计有4 292 000例癌症新发病例, 2 814 000例癌症患者死亡, 也就意味着全国每一分钟就有8人患癌, 5人因癌症死亡, 恶性肿瘤已成为我国最主要的致死疾病之一。
目前研究认为活性氧的作用导致的DNA氧化损伤是人类癌症发生的一个重要因素, 而癌症的发生需要满足以下两个条件:一是DNA必须受到了一定的损害后使细胞失去了本身的生长调节机制(这个过程被称为肿瘤的始动) ;第二是由于炎症瀑布反应进一步导致正常免疫细胞识别肿瘤细胞能力的丧失。最后, 活性氧类物质干扰了程序性细胞死亡的基本细胞控制即细胞凋亡。许多临床研究发现, 肿瘤疾病患者的血清中Co Q10水平要较正常人的水平要低且具有统计学意义, 因此怀疑Co Q10与肿瘤的发生发展有关。
Co Q10最早于1948年被发现, 由于在人体器官中存在以及在生理等方面的重要功能, 又被称为维生素Q或维生素辅酶Q。1957年Crane等在牛心中首次分离出Co Q10。1978年, Peter Mitchell发现了Co Q10在生物能量转移方面起重要作用, 并因此获得了诺贝尔奖。Co Q10是生物醌化合物, 以多种形式广泛存在于包括酵母, 植物、动物等生物体中。目前Co Q10在人类和动物中是发现和研究最多的, 它主要有两种生理活性, 一是参与线粒体中呼吸链的氧化反应增强线粒体中ATP的合成;二是转变成具有抗氧化活性的还原物(泛醇) 参与抗氧化作用。因此Co Q10现被用作膳食补充剂和临床疾病治疗, 临床上现已将Co Q10用于原发性Co Q10缺乏症、线粒体病、心血管病、神经系统疾病、肿瘤等疾病的治疗。Crane等证实了Co Q10是线粒体呼吸链上的成员并承担了重要的作用, 它参与了细胞代谢, 作为线粒体内外膜的电子载体, 还防止了膜蛋白上的蛋白质氧化和脂质的过氧化。同时, Co Q10的抗氧化作用可以阻止损伤细胞的自由基和活性氧, 通过氧化还原信号来控制细胞的能量代谢和死亡的调节。许多化疗药物有清除氧自由基作用, 如阿霉素、足叶乙甙、甲氨蝶呤、喜树碱等, 这些抗癌治疗药物在癌细胞中通过诱导提高癌细胞中的Co Q10水平, 促进抗氧化作用来减少自由基产生, 因此将Co Q10用于临床治疗可成为抗癌症治疗的一个重要手段。以下是关于临床上Co Q10治疗癌症患者的研究现状。
5.2 辅酶Q10在相关肿瘤治疗中的应用
5.2.1 乳腺癌
乳腺癌在中国女性恶性肿瘤中位于第一位, 每年新发病例约27.9万。而Co Q10在肿瘤当中关于乳腺癌的临床研究是最多的, 多个临床研究发现女性乳腺癌患者血浆和乳腺癌组织中Co Q10水平明显下降, 且水平的高低与患者的预后相关。Aponte等关于Co Q10在乳腺癌临床治疗作用的系统评价中共纳入了6篇报告, 研究者让实验组患者每日补充100 mg的Co Q10, 结果提示Co Q10可以增加乳腺癌患者机体抗氧化物质的水平, 降低血浆细胞因子及肿瘤标志物水平。Sachdanandam等的研究将实验组乳腺癌患者分为两组, 一组为“100 mg的Co Q10与他莫昔芬10 mg, 2次/d, 共45 d”, 另一组时间延长至90 d。实验结果示Co Q10可以通过提高基质金属组织蛋白酶抑制剂1、2(TIMF1、TIMF 2) 的水平来降低基质金属蛋白酶-2、9(MMP-2、MMP-9)(P<0.05) , 还可以降低血浆细胞因子如白细胞介素(IL1β、IL6、IL8) 、肿瘤坏死因子α(TNF-α) 、血管内皮生长因子(VEGF) , 以及肿瘤标志物水平如癌胚抗原(CEA) 、糖类抗原15-3(CA15-3) , P>0.05, Panchanatham等的实验结果证明Co Q10联合他莫昔芬可增强乳腺癌患者化疗效果, 该研究提示了Co Q10可能可作为乳腺癌的联合治疗方案之一。但Chai等的报道中提出了不同的观点, 他们的研究发现绝经后乳腺癌患者的血浆Co Q10水平与乳腺癌风险呈正相关(P=0.04) , 这提示了绝经后妇女高Co Q10水平可能增加女性患乳腺癌的风险。未来需要更大样本、多中心的前瞻性研究, 更长的随访时间以及对循环Co Q10的生理调节和功能的进一步研究, 来确定Co Q10在乳腺癌中的潜在作用。
5.2.2 黑色素瘤
Persaud等的研究中, 给予黑色素瘤小鼠模型Co Q10脂质体外用制剂共30 d, 与对照组相比, 治疗组小鼠的肿瘤体积较对照组平均减少53.25%, 且治疗组的组织学分析显示肿瘤血管系统的结构完整性较对照组显著降低。Narain等的实验中分析了Co Q10对黑色素瘤中bcl-2基因表达的影响, 实验组模型用50μmol/L Co Q10处理24 h后, 肿瘤细胞Livin及Survivin基因的表达和bcl-2蛋白水平较对照组显著下调(P<0.05) ;而在新生纤维母细胞中的bcl-2水平并没有明显改变, 且Livin及Survivin基因的表达是增加的。笔者认为Co Q10通过促使黑素瘤细胞凋亡潜能的恢复来调节bcl-2水平和存活蛋白的表达, 是一种不会负面影响正常皮肤细胞代谢的方式, 该实验提示了Co Q10可能通过靶向诱导bcl-2蛋白家族来起到抗癌的作用。Rusciani等的研究中纳入了117例被临床或影像学证明没有发生转移的黑色素瘤患者, 在肿瘤的发展过程中, 分别对比患者在T1、T2、T3、T4(TMN分期) 阶段血清Co Q10的水平, 最后发现发生转移的患者比无转移亚组患者的Co Q10水平显著降低, 这提示了黑色素瘤患者血清中Co Q10的基线水平可能是一个可用于评估黑素瘤进展风险的孤立且强大的预后因子, 但仍需要更大的多中心的临床随机对照试验进一步明确。在Rusciani等的另一个研究中, 利用重组干扰素α-2b联合Co Q10治疗恶性黑色素瘤患者, 通过评估5年中治疗患者的复发率来判断治疗效果。所有患者均完成治疗及随访, 结果表明重组干扰素α-2b联合Co Q10与对照组相比可以显著降低恶性黑色素瘤的复发率, 而且Co Q10的副作用很少, 使其可作为Ⅱ期黑色素瘤术后新的辅助治疗方式。但是由于作者采集的数据质量不高, 且由于样本量小以及非随机对照试验导致存在了结果存在很大的偏移。
5.2.3 肝癌
Fouad等在肝细胞癌的治疗研究中, 给予肝癌大鼠模型实验组Co Q10每天0.5 g/kg的剂量共四周, 最后观察大鼠生化参数丙二醛(MDA) , 肿瘤坏死因子(TNF) 和一氧化氮(NO) 的水平, 以及大鼠肝脏组织病理学和免疫组织化学结果。生化参数提示Co Q10显著抑制了脂质过氧化, 防止还原型谷胱甘肽耗竭和超氧化物歧化酶活性的降低, 同时降低了肝组织细胞中肿瘤坏死因子水平及阻止了一氧化氮的升高;免疫组化提示Co Q10可明显降低大鼠肝癌细胞中的Hep Par-1、甲胎蛋白、诱导型一氧化氮合酶、环氧合酶-2及κB核因子的表达(P<0.05) , 提示Co Q10可能是肝癌的一个潜在治疗药物。国内一课题组Liu等测定了71例原发性肝癌患者手术前后的Co Q10水平, 结果显示肝癌患者手术前后的Co Q10水平均较正常参考值低, 且术后的Co Q10水平较术前显著下降(P<0.01) , 研究者认为通过外源性补充Co Q10来纠正术后Co Q10的缺乏可改善肝癌患者手术的预后。Liu等设计了Co Q10应用于肝细胞癌患者手术后的随机对照试验, 其中实验组的21例肝癌患者给予300 mg/d的Co Q10治疗量, 最终证实了Co Q10可以提高肝癌患者术后的抗氧化能力, 及减少机体内的氧化应激及炎症水平。
5.2.4 结肠癌
目前尚未检索到关于Co Q10在结肠癌患者身上的临床研究。变异腺窝病灶(ACF) 及黏蛋白缺乏病灶(MDF) 被认为是大肠的癌前病变, Sakano等通过在大鼠模型体内注射“氧化偶氮甲烷”来诱发大鼠结肠产生ACF和MDF, 实验组大鼠同时给予服用Co Q10共4周, 结果提示实验组大鼠使用Co Q10治疗后ACF及MDF较对照组明显减少(P<0.05) , 笔者认为Co Q10可能有预防结肠癌变的作用。Kim等在大鼠体内注射二甲肼(DMH) , 导致大鼠白细胞DNA损伤以及诱导环氧合酶-2(COX-2) 基因和诱导型一氧化氮合酶(i NOS) 基因的表达, 使大鼠模型产生ACF, 实验结果显示同时给予Co Q10喂养的实验组大鼠的ACF较对照组降低20%(P<0.05) , 而且实验组大鼠体内COX-2和i NOS水平较对照组降低(P<0.05) , 推测Co Q10可能通过调整结肠黏膜细胞中COX-2和i NOS的基因表达和白细胞的DNA损伤的机制, 来阻止结肠癌前病变的恶化及阻止结肠发生癌变。
5.2.5 其他肿瘤与辅酶Q10的相关性
Chai等统计了307例前列腺癌患者血清Co Q10水平与前列腺患病危险因素, 最终结果显示患前列腺癌与Co Q10的水平无明显相关性(P=0.50) , 但由于跟踪年限过短及晚期前列腺癌患者的病例数过少, 因此需要做更进一步的研究证实。在肺癌与Co Q10相关性研究中, Co Q10治疗组肺癌患者代表脂质过氧化作用的MDA水平及DNA损伤率较对照组明显降低(P<0.001) 。宫颈上皮内瘤变和宫颈癌患者血浆Co Q10的水平降低也被证实, 但目前还没有相关的临床试验报道。
5.3 辅酶Q10临床治疗的方案及安全性
在一篇系统评价中认为Co Q10在癌症治疗过程中可以减低蒽环类药物对心脏及肝脏毒性作用, 这使得Co Q10可能成为化疗方案中一种新的辅助治疗方式。目前的研究及临床应用来看, 即使在理论上的最高剂量, Co Q10用于临床治疗也是安全的, 临床试验中还没有发现有因Co Q10引起的严重的不良反应需要停止对患者治疗的报告, 但是, Co Q10的治疗可能会发生如腹部不适、恶心、呕吐、腹泻、厌食症等症状, 但大多很轻微, 过敏性皮疹和头痛也偶有报道。此外, Co Q10的抗血小板作用可能会增加出血的风险。Co Q10是通过肝脏进行生物转化, 主要通过经胆道清除, 因此它可以积聚在肝损害或胆道梗阻的患者体内, 进一步加重肝脏负担。Co Q10对于癌症治疗的效果及能否改善患者预后仍需得到进一步充分的评估, 因此仍需大型的多中心前瞻性试验进一步来证实Co Q10对于癌症的治疗效果、安全性及临床上最佳的治疗剂量。
六、辅酶Q10的抗氧化与抗疲劳作用及临床应用
6.1 目前国内外市场上已有的辅酶Q10补充剂
市场上已有的辅酶Q10补充剂主要应用于医药行业, 目前辅酶Q10的针剂、胶囊、软胶囊、片剂都有生产, 另外, 其在食品领域常作为各类营养保健食品添加剂被广泛应用。除此之外, 采用新型纳米技术制备的辅酶Q10纳米脂质体因表现出较高的生物利用度而受到欢迎。
6.2 辅酶Q10的抗氧化作用
辅酶Q10存在于线粒体内膜上, 与细胞产能过程密切相关, 在细胞呼吸和ATP产能过程中起到传递电子和转运蛋白的作用。辅酶Q10在线粒体中主要以两种形式存在:氧化型和还原型。氧化型可以通过丢失两个电子转化成还原型, 并以还原型形式起到抗氧化作用。还原型辅酶Q10还可以抑制自由基介导的膜脂蛋白氧化损伤, 清除游离自由基、抑制氧化应激反应, 从而起到抗氧化功能。
动物实验表明, 通过将48只雄性老龄SD大鼠按其血清丙二醛含量分层后随机分为3个剂量试验组和1个阴性对照组, 每组12只大鼠。3个剂量组分别给予15.00、7.50、3.75mg/kg剂量的辅酶Q10, 阴性对照组给予小麦胚芽油。采用经口灌胃法给样, 每日1次, 连续灌胃90天。试验终末, 用试剂框测定大鼠体内丙二醛、脂褐质的含量以及超氧化物歧化酶、谷胱甘肽过氧化物酶的活性。结果显示:与阴性对照组相比, 辅酶Q10各剂量组中的大鼠血清、脑和肝组织中的丙二醛含量呈不同程度下降;超氧化物歧化酶和谷胱甘肽过氧化物酶的活性呈不同程度升高;脑和肝组织中脂褐质有不同程度下降。说明辅酶Q10对大鼠有抗氧化作用。
人体试验表明, 120名健康受试者按血清中丙二醛含量随机分为样品组和安慰剂组, 样品组连续服用辅酶Q10胶囊120天, 每日2次, 每次1粒, 胶囊内含辅酶Q1020mg, 通过检测受试者血清中丙二醛含量及超氧化物歧化酶、谷胱甘肽过氧化物酶的活性以及安全性指标来评价其抗氧化功能。试验结果表明:样品组中血清内丙二醛含量下降率、超氧化物歧化酶和谷胱甘肽过氧化物酶活性升高率均显著大于安慰剂组(P<0.01) , 说明辅酶Q10对人体具有抗氧化作用, 而两组人群中的各项安全性指标试验均无明显改变, 说明辅酶Q10长期使用表现出较好的安全性。除此之外, 在多种疾病的辅助治疗中也显示, 辅酶Q10可通过产生超氧化物阴离子自由基来清除游离自由基、抑制氧化应激反应, 表现出抗氧化作用。
6.3 辅酶Q10的抗疲劳作用
辅酶Q10另一个非常显著的优点, 就是兼具缓解体力疲劳的功能。随着年龄增加, 人体内的辅酶Q10含量逐渐减少, 细胞产能也受到影响, 补充外源性的辅酶Q10可以保障细胞呼吸链的通畅, 增加机体肝糖原的储备。因此, 辅酶Q10可以通过抗氧化和改善疲劳两个方面同时“抗衰老”, 这是其他抗氧化剂无可比拟的优势。
动物实验表明, 分别以0、1.5、15、45mg/kg剂量的辅酶Q10给小鼠灌胃, 每日1次, 连续给药4周, 通过负重游泳试验, 测定运动后的血清尿素氮含量以及肝糖原储备来评价其抗疲劳功能。试验结果表明, 辅酶Q10能够显著延长小鼠的负重游泳时间(15mg/kg, P>0.05) , 增加肝糖原储备(15mg/kg, P>0.01;45mg/kg, P>0.05) , 减少小鼠运动后血清尿素氮含量(1.5, 15, 45mg/kg, P>0.01) , 提示辅酶Q10可以增强小鼠的游泳耐力并提高其抗疲劳能力。
Mizuno K等对17名健康志愿者随机分为口服辅酶Q10组(每天100或300mg) 和安慰剂组, 给药8天。在自行车测功计(bicycle ergometer) 上进行超负荷运动试验, 通过检测最大速度变化情况以及视觉类似物量表(visual analog scale, VAS) 评价其抗疲劳作用。结果表明, 300mg辅酶Q10的试验组与安慰剂组比较, 在超负荷试验中, 受试者最大速度变化率较高, 试验结束后受试者主观疲劳感较轻且所需恢复时间较短。结果表明:长时间运动时可明显引起线粒体膜磷脂的减少, 而口服辅酶Q10补充剂具有防止线粒体膜磷脂减少的保护效果, 并能提高肌肉中磷酸肌酸从运动中恢复的能力。因此, 使用辅酶Q10可通过抗氧化作用减轻身体疲劳感并有助于体内氧化性磷酸化过程。但在此试验中, 100mg辅酶Q10的试验组与安慰剂组相比, 各项指标均无明显差别, 说明辅酶Q10只有在充足剂量下才能发挥抗疲劳作用。
6.4 辅酶Q10的抗氧化与抗疲劳作用在临床方面的应用
6.4.1 在癌症方面的应用
癌症相关性的疲劳在各种形式和阶段的癌症患者中是最为常见的症状之一, 但临床关注较少。癌症相关性的疲劳与细胞氧化应激有关。在癌症治疗期间, 药物过量诱导的氧化应激可以限制治疗效果并带来各种不良作用, 包括疲劳、恶心、呕吐等。Nicolson GL就近年来对脂质替代疗法在癌症疾病的应用进行了综述, 指出脂替代疗法(膜脂与抗氧化剂及辅酶因子, 如辅酶Q10, 联合使用) 可以减少癌症患者的细胞膜氧化损伤, 恢复线粒体电子传递功能和其他细胞功能, 并在不降低治疗结果的前提下改善慢性疲劳、减轻不良反应。
另外, 辅酶Q10还具有潜在的治疗癌症的能力并用于辅助治疗。动物实验表明, 患有肝细胞癌的大鼠通过腹腔注射辅酶Q100.5mg/kg, 每日1次, 连续注射4周, 检测大鼠肝脏中还原性谷胱甘肽和超氧化物歧化酶活性、肿瘤坏死因子α和一氧化氮(NO) 含量等指标来评价辅酶Q10对氧化应激的作用及其改善癌症的效果。试验结果表明, 辅酶Q10能显著抑制脂质过氧化, 防止还原性谷胱甘肽和超氧化物歧化酶活性的降低, 减少大鼠肝脏组织中肿瘤坏死因子α和NO的含量, 改善不良病理变化。说明辅酶Q10对于治疗肝癌具有潜在的能力。
Hertz N等对41名晚期癌症患者进行长达9年的试验研究, 给予癌症患者每日300mg剂量的辅酶Q10以及其他剂量的抗氧化剂(维生素C, 硒, 叶酸等) 作为癌症治疗的补充剂, 这些抗氧化剂每日分2次给予, 在给药2个月后通过Kaplan–Meier曲线预测患者存活时间。试验结果表明:10名患者(24%) 实际生存时间短于预期值, 31名患者(76%) 实际生存时间长于预期值;治疗期间, 患者对不良反应耐受较好。
6.4.2 在心脏疾病方面的应用
冠心病患者心脏组织中辅酶Q10的含量明显低于正常人群, 并且血管壁的超氧化物歧化酶活性降低。内皮细胞结合的超氧化物歧化酶与内膜依赖性舒张有关, 而改善患者内膜依赖性舒张有利于缓解病症。Tiano L等对19名冠心病患者给予辅酶Q10口服制剂, 300mg每天, 给药1个月后, 所有患者均进行肱动脉内皮依赖性评估、心肺运动试验和内皮细胞结合的超氧化物歧化酶活性的测定。结果显示:与安慰剂组相比, 辅酶Q10组患者细胞外超氧化物歧化酶活性、内膜依赖性舒张以及最大摄氧量均显著升高。Lee BJ等进行的试验表明, 给予37名冠心病患者辅酶Q10补充剂(60或150mg/d) , 持续12周, 每4周测定患者血浆中辅酶Q10浓度、丙二醛的含量及抗氧化酶活性。结果显示:150mg辅酶Q10组患者血浆辅酶Q10浓度较安慰剂组显著升高(P<0.001) 。给药4周及8周后, 丙二醛浓度显著降低(P=0.03) ;给药12周后, 过氧化氢酶(P=0.03) 及超氧化物歧化酶(P=0.03) 活性显著升高, 患者氧化应激反应被抑制, 并且较高剂量的辅酶Q10补充剂(>150mg/d) 可能会加速并维持冠心病患者的抗氧化。
6.4.3 在纤维肌痛方面的应用
纤维肌痛(FM) 是以慢性、广泛的关节和肌肉疼痛为特征, 是病因不明的综合征。Miyamae T等通过对10名FM患者和67名健康受试者进行对照研究, 发现FM会引起体内氧化应激增强以及辅酶Q10缺失。之后, 给予FM病人还原性辅酶Q10, 100mg/d, 持续12周, 观察患者血浆中还原性辅酶Q10浓度、胆固醇脂及游离脂肪酸的浓度来评价辅酶Q10对改善FM症状的疗效。结果显示:患者体内辅酶Q10浓度增加, 应激反应被改善, 游离胆固醇和胆固醇脂水平下降。表明辅酶Q10可加快胆固醇代谢并改善肝功能。在试验中, 还原性辅酶Q10对生命质量(QOL) 的影响也进行了评估, Chalder疲劳量表显示辅酶Q10可改善慢性疲劳, 说明该补充剂对改善FM临床症状有潜在的疗效。Cordero MD等进行的试验也表明, FM与线粒体功能障碍及氧化应激反应有关, 分别给予20名FM患者和10名健康受试者辅酶Q10口服制剂, 每天300mg, 分3次给予, 连续给予3个月, 使用纤维肌痛影响问卷(FIQ) 、视觉类似物量表(VAS) 和头痛冲击试验(HIT-6) 来评价临床疗效。结果显示:服用辅酶Q10后, 患者头疼、疲劳等临床参数, 显著改善(P<0.001) 。
七、辅酶Q10在免疫调节中的作用
研究辅酶Q10对小鼠免疫调节功能的影响, 并初步探讨其作用机制。方法:从细胞免疫、体液免疫、单核-巨噬细胞吞噬作用及自然杀伤细胞的攻击作用四方面观察辅酶Q10对小鼠免疫功能的影响。结果:与阴性对照组(蒸馏水) 和溶剂对照组(植物油) 比较, 辅酶Q10 8.0、12.0和24.0 mg/kg.BW剂量组均能提高ConA诱导的小鼠脾淋巴细胞的增殖能力;4.0、8.0和12.0 mg/kg.BW剂量组均能增加小鼠左后足跖部厚度差24 h测量值;12.0和24.0 mg/kg.BW剂量组均能提高小鼠溶血空斑数;4.0、8.0、12.0和24.0 mg/kg.BW剂量组均能提高小鼠NK细胞活性及小鼠腹腔巨噬细胞吞噬鸡红细胞的吞噬率;12.0 mg/kg.BW剂量组能提高小鼠腹腔巨噬细胞吞噬鸡红细胞的吞噬指数。结论:辅酶Q10具有增强小鼠免疫力作用, 其作用机制可能与辅酶Q10能激活NK细胞、T细胞、巨噬细胞功能及清除氧自由基、稳定膜电位等有关。
由于辅酶Q10具有醌式及异戊烯基侧链的结构特点, 决定了它在生物体中还具有许多重要生理功能。近年来, 越来越多的研究发现还原型辅酶Q10(辅酶Q10 H2, 辅酶q10醇) 具有抑制自由基介导的膜脂蛋白氧化损伤作用, 因此, 还原型辅酶Q10的抗氧化作用, 及其在氧化应激相关疾病的预防和治疗中的作用正受到越来越多的关注。通过从动植物提取、化学合成、微生物发酵的方式得到的辅酶Q10制剂作为能量合成物质已广泛应用于临床, 目前辅酶Q10主要用于防治急性心肌缺血时心肌收缩力减弱、降低血压、增加心输出量等方面。迄今为止, 辅酶Q10临床与实验研究主要集中于若干器官和疾病时线粒体的形态、功能和代谢的变化, 如心脏病、糖尿病等, 其对免疫系统的作用以及在肿瘤治疗中辅助作用的研究报道较少。本文通过对辅酶Q10在小鼠体内免疫调节作用的研究, 旨在说明其对细胞免疫、体液免疫、单核-巨噬细胞吞噬及自然杀伤细胞系统的调节作用, 为辅酶Q10用于免疫功能低下疾病的治疗、辅助治疗及人群的日常保健提供实验依据。
7.1 材料与方法
7.1.1 材料
7.1.1.1 实验动物
清洁级ICR雌性小鼠, 体重18~22 g, 由中科院上海实验动物中心提供, 实验动物生产许可证号为SCXK(沪) 2002-0010。
7.1.1.2 受试物
辅酶Q10由浙江省生产辅酶Q10的药业有限公司提供, 批号为20051021。
7.1.2 方法
参照《免疫毒理学实验技术》进行。试验项目包括:
7.1.2.1 剂量设计
实验设5个剂量组、1个阴性对照组和1个溶剂对照组。剂量分别为2.0、4.0、8.0、12.0、24.0 mg/kg·BW。
7.1.2.2 小鼠脏器/体重比值测定(包括胸腺/体重和脾脏/体重比值)
雌性ICR小鼠70只, 体重18~22 g, 随机分7组。每日分别以2.0、4.0、8.0、12.0、24.0 mg/kg·BW 剂量给予受试物, 连续灌胃给予受试物 30 d。另设阴性对照组和溶剂对照组。
7.1.2.3 细胞免疫功能测定
7.1.2.3.1 ConA诱导的小鼠脾淋巴细胞转化试验(MTT) 法:动物及剂量同1.2.2。
7.1.2.3.2 绵羊红细胞(SRBC) 诱导小鼠DTH(足跖增厚法) : 动物及剂量同1.2.2。
7.1.2.4 体液免疫功能测定
7.1.2.4.1 小鼠血清溶血素测定(血凝法) : 动物及剂量同1.2.2。
7.1.2.4.2 抗体生成细胞检测(Jerne改良玻片法) : 动物及剂量同1.2.2。
7.1.2.5 单核巨噬细胞吞噬功能测定
7.1.2.5.1 小鼠腹腔巨噬细胞吞噬鸡红细胞试验(半体内法) : 动物及剂量同1.2.2。
7.1.2.5.2 小鼠碳廓清试验: 动物及剂量同1.2.2。
7.1.2.6 NK细胞活性测定(LDH测定法) 动物及剂量同1.2.2。
7.1.2.7 统计方法
全部数据采用SPSS/PC软件包在微机上进行统计处理。
7.2 结果
7.2.1 对小鼠免疫器官的影响
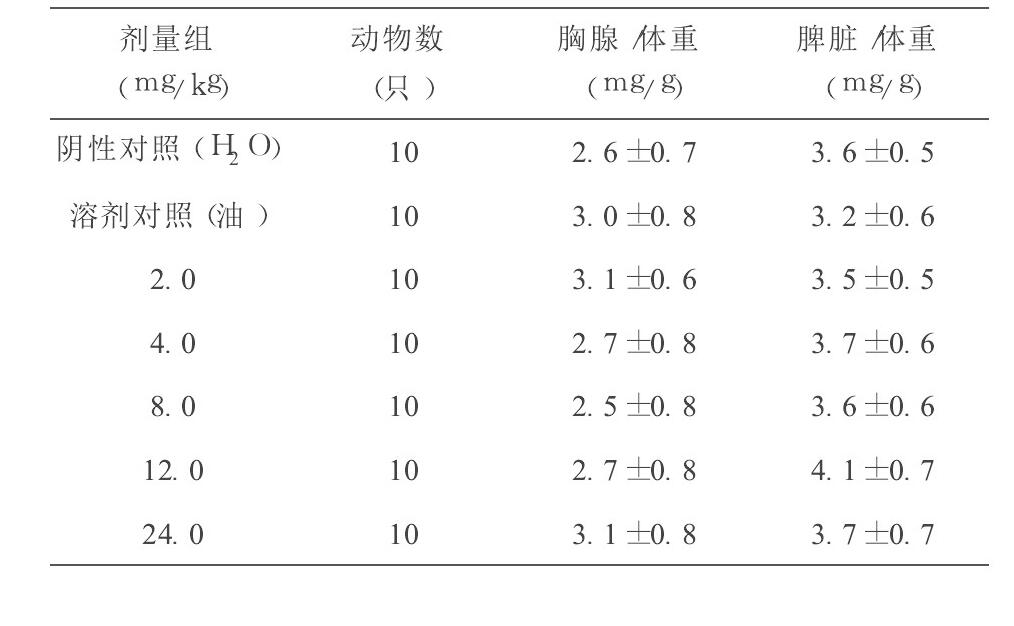
从表1可见, 辅酶Q10各剂量组小鼠胸腺/体重、脾脏/体重比值和阴性对照组(蒸馏水) 、溶剂对照组(植物油) 比较, 差异均无显著性(方差分析, P>0.05) 。
7.2.2 对小鼠细胞免疫功能的影响
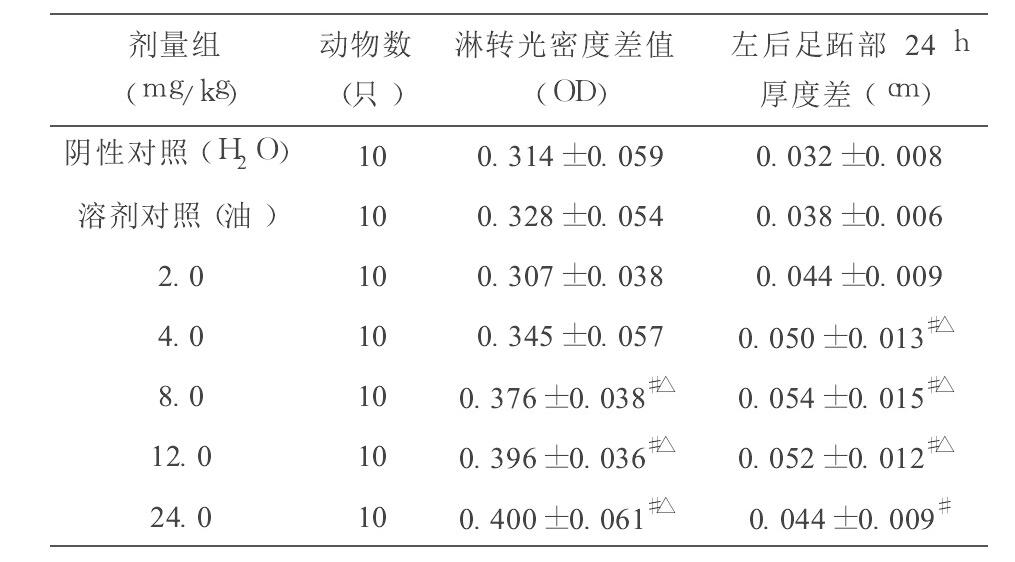
从表2可见, 与阴性对照组(蒸馏水) 、溶剂对照组(植物油) 比较, 辅酶Q10 8.0、12.0和 24.0 mg/kg·BW 剂量组均能提高ConA诱导的小鼠脾淋巴细胞的增殖能力, 4.0、8.0和 12.0 mg/kg·BW 剂量组均能增加小鼠左后足跖部厚度差 24 h 测量值, 差异均有显著性(q检验, P>0.05) ;与阴性对照组(蒸馏水) 比较, 辅酶Q10 24.0 mg/kg·BW 剂量组能增加小鼠左后足跖部厚度差 24 h 测量值, 差异有显著性(q检验, P>0.05) , 但和溶剂对照组(植物油) 比较, 差异无显著性(q检验, P>0.05) 。
7.2.3 对小鼠体液免疫功能的影响
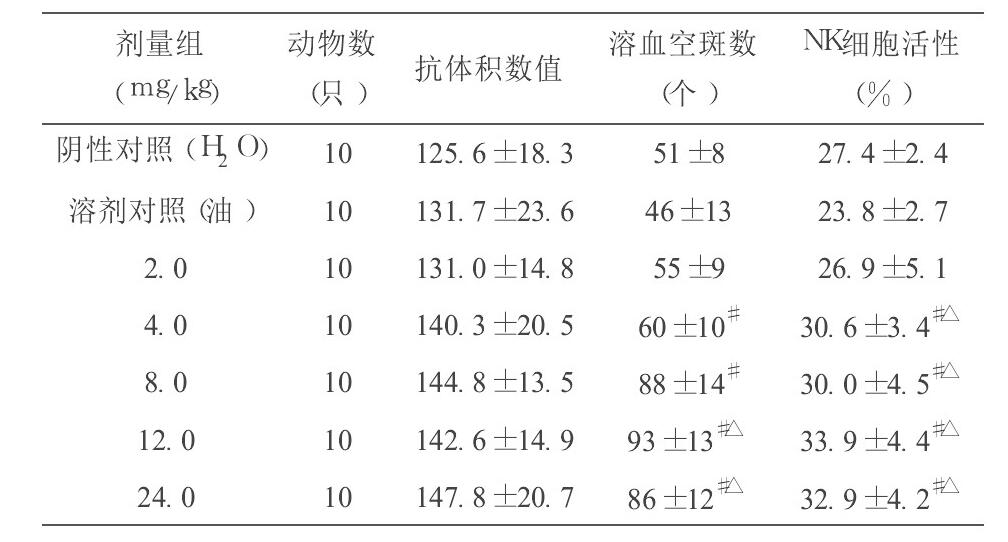
从表3可见, 与阴性对照组(蒸馏水) 、溶剂对照组(植物油) 比较, 辅酶Q10 200.0和 400.0 mg/kg·BW 剂量组均能提高小鼠溶血空斑数, 差异均有显著性(q检验, P>0.05) ;与阴性对照组(蒸馏水) 比较, 辅酶Q10 4.0和 8.0 mg/kg·BW 剂量组能增加小鼠溶血空斑数, 差异有显著性(q检验, P>0.05) , 但和溶剂对照组(植物油) 比较, 差异无显著性(q检验, P>0.05) 。
7.2.4 对小鼠单核巨噬细胞吞噬功能的影响
从表4可见, 与阴性对照组(蒸馏水) 比较, 辅酶Q10 4.0、8.0、12.0和 24.0 mg/kg·BW 剂量组均能提高小鼠腹腔巨噬细胞吞噬鸡红细胞的吞噬率, 差异均有显著性(q检验, P>0.05) 。12.0 mg/kg·BW 剂量组能提高小鼠腹腔巨噬细胞吞噬鸡红细胞的吞噬指数, 差异有显著性(q检验, P>0.05) 。8.0 mg/kg·BW 剂量组小鼠腹腔巨噬细胞吞噬鸡红细胞的吞噬指数和溶剂对照组(植物油) 比较, 差异有显著性(q检验, P>0.05) , 但和阴性对照组(蒸馏水) 比较, 差异无显著性(方差分析, P>0.05)
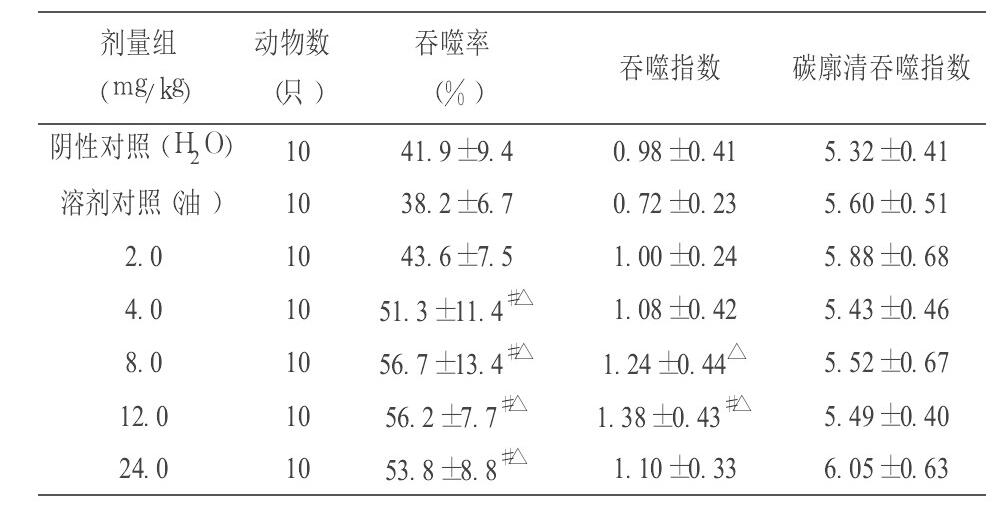
7.2.5 对小鼠NK细胞活性的影响
从表3可见, 与阴性对照组(蒸馏水) 比较, 辅酶Q10 66.7、133.3、200.0和 400.0 mg/kg·BW 剂量组均能提高小鼠NK细胞活性, 差异均有显著性(q检验, P>0.05) 。
7.3 讨论
国内外实验和临床研究表明, 辅酶Q10对心脏病、高血压、脑血管障碍、坏血病、动脉粥样硬化等疾病有较好的疗效, 可作为肌体细胞代谢及细胞呼吸激活剂。辅酶Q10也是一种机体的非特异性免疫增强剂, 它能增强机体免疫细胞杀死细菌的活力, 增加白细胞的数量、胸腺活力、免疫球蛋白含量和抗体的量。
从本试验结果可见, 不同剂量的辅酶Q10对小鼠主要免疫器官(胸腺和脾脏) 无明显增强或抑制作用。辅酶Q10剂量为8.0、12.0和 24.0 mg/kg·BW 时能提高ConA诱导的小鼠脾淋巴细胞的增殖能力, 剂量为4.0、8.0和 12.0 mg/kg·BW 时能增加小鼠左后足跖部厚度差 24 h 测量值, 与对照组比较均有显著性差异, 说明辅酶Q10能提高小鼠细胞免疫功能; 辅酶Q10 12.0和 24.0 mg/kg·BW 剂量组均能提高小鼠溶血空斑数, 与对照组比较有显著性差异, 说明辅酶Q10对小鼠体液免疫具有一定的调节作用; 辅酶Q10 4.0、8.0、12.0和 24.0 mg/kg·BW 剂量组均能提高小鼠腹腔巨噬细胞吞噬鸡红细胞的吞噬率, 12.0 mg/kg·BW 剂量组能提高小鼠腹腔巨噬细胞吞噬鸡红细胞的吞噬指数, 与对照组比较均有显著性差异, 说明辅酶Q10能增强单核- 巨噬细胞吞噬功能;辅酶Q10 4.0、8.0、12.0和 24.0 mg/kg·BW 剂量组均能提高小鼠NK细胞活性, 与对照组比较有显著性差异, 说明辅酶Q10能增强小鼠NK细胞活性。
参与免疫调节的细胞主要有T细胞、B细胞、NK细胞和巨噬细胞。T细胞作为免疫效应细胞主要行使两方面功能, 即作为TDTH介导DTH反应和作为Tc细胞直接杀伤靶细胞, T细胞又是免疫调节细胞, 具有辅助其他免疫细胞分化和调节免疫应答的功能, 因此, T细胞在机体的细胞免疫和体液免疫诱导中均有重要作用。Mφ是体内非常重要的免疫细胞, 在抗感染免疫和肿瘤免疫等方面都起着重要的作用, 可通过处理抗原和释放可溶性因子对免疫功能起重要的调节作用。NK细胞参与非特异性免疫, 是机体天然免疫的主要承担者, 同时它对T细胞、B细胞、骨髓干细胞等均有调节作用, 并通过释放细胞因子对机体免疫功能进行调节。从我们的实验结果可见, 辅酶Q10能提高ConA诱导的小鼠脾淋巴细胞的增殖能力, 增强小鼠迟发型变态反应, 激活单核-巨噬细胞吞噬系统, 提高NK细胞活性。由此我们可以认为辅酶Q10对小鼠免疫功能的调节作用可能主要是通过激活NK细胞活性, 从而启动T细胞的功能, 提高T细胞的应答能力, 使巨噬细胞活化成为抗原提呈细胞, 参与识别、吞噬和处理抗原、传递免疫信息等免疫反应过程来实现的。
机体的免疫反应包含了特异性免疫反应和非特异性免疫反应。从本实验结果可见, 辅酶Q10不仅能增强非特异性免疫反应, 同时也能增强特异性免疫反应。
大量的研究结果已证实, 辅酶Q10具有抗氧化作用和自由基清除功能, 可减少机体组织的氧化损伤。因此, 从另一方面来讲, 辅酶Q10增强小鼠免疫功能可能和其清除氧自由基, 从而减少了自由基对免疫细胞的损伤, 促进细胞膜结构的完整性, 稳定膜电位等有关。
八、辅酶Q10的开发
8.1 工艺开发
辅酶Q10的生产主要有3种方法:一是动植物组织提取法;二是化学合成法;三是微生物发酵提取法。其中合成法又分为全合成法和半合成法, 目前研究的基本都是半合成法;发酵法包括直接生物发酵法和前体微生物生物转化法。
直接提取法主要是从动物新鲜肝脏中提取, 因为天然含量低, 各种化学成份复杂, 因而生产成本高, 生产规模受到限制。
8.1.1 提取法
主体工艺路线如下:
将原料进行处理, 一般采用碱性乙醇进行均质皂化, 石油醚等溶剂进行萃取, 减压浓缩, 并用硅胶等柱层析进行分离, 最后在乙醇中低温进行结晶。
利用猪心残渣生产辅酶Q10的配料比为猪心残渣∶95%乙醇∶氢氧化钠∶焦性没食子酸=1∶1.3∶0.13∶0.12。
在反应罐中加入乙醇氢氧化钠和焦性没食子酸, 搅拌后加入提取细胞色素后的猪心残渣, 在78~90℃下回流皂化30min, 冷至35℃, 得皂化液。
萃取液的配料比为皂化液∶石油醚=1∶1.4。
在上述皂化液中加入0.6倍量的石油醚, 搅拌提取, 静止分层后, 吸出上清液, 残渣再分别用0.4倍量的石油醚萃取2次, 合并萃取液, 水洗直至至近中性。
将萃取液在40℃以下减压浓缩至原体积的1/20, 冷却、静置后过滤, 得浓缩滤液。
在长径比等于7∶1的纯化柱中装好硅胶, 将上述猪心残渣∶硅胶(60~100目) =1∶0.01浓缩滤液通过硅胶进行吸附, 吸附后先用石油醚或120#汽油洗柱除去杂质, 待流出液为五无时, 再用含5%乙醚的石油醚洗脱, 至洗脱液近乎无色, 得洗脱液。
将洗脱液在40℃以下减压浓缩, 除去石油醚后得棕色油状物。于油状物中加入热的无水乙醇, 溶解后冷却结晶11h, 滤干, 真空干燥, 得辅酶Q10。上述工艺能够从每kg猪心中提取到50~70mg辅酶Q10。此法工艺简单, 是经典的提取辅酶Q10的方法。改进提取工艺, 可采用碱水替代碱性乙醇进行皂化, 使用复合溶剂提取或不经皂化提取, 但从技术发展、市场需求的角度来看, 特别是从辅酶Q10的应用渠道不断拓宽使市场需求持续增长的状况来看, 提取法将逐渐被大规模的合成法或发酵法所替代。
| 物种 | 基因种类 |
|---|---|
| 大肠杆菌 | IspA, IspG, UbiE, UbiH, ubiF, ubiB, ubiC, ubiD/ubiX |
| 酿酒酵母 | COQ2, COQ3, COQ4, COQ5, COQ6, COQ7, COQ8 |
| 粟裂酵母 | Ppt1, SPCC162.05, SPAC1687.12c, SPCC4G3.04c, SPBC146.12, SPBC337.15c, Abc1 |
| 线虫 | F57B9.4b, CeCOQ3, TO3F12, ZK652.9, K07B1.2, Clk-1, C35D10.4 |
| 人类 | HuPPT1, HuCOQ3, LOC51117, MGC4767, MGC23201, HuCOQ7, AAH13114 |
| 拟兰芥 | AtPPT1, AtCOQ3, At2g03690, At5g57300, At3g24200, AtABC1 |
8.1.2 茄尼醇法
辅酶Q10的半化学合成法采用昂贵的茄尼醇为原料。茄尼醇这种天然的长链半萜烯醇化合物, 结构独特(异戊二烯九聚体, 双键全反式构型) , 完全类似于辅酶Q10的支链, 以它合成辅酶Q10可以避免复杂冗长的逐步延长侧链的过程, 同时在保持双键几何构型和立体、区域选择性上有着不可替代的优势。我国是全球最大烟草生产国, 有得天独厚的原料资源优势。茄尼醇不仅可以直接从烟草中提取, 而且还可从废次烟叶中获得, 这更降低了原料成本, 也实现了变废为宝, 提高了烟草废弃物的资源再生和综合利用率。因此, 用茄尼醇为原料半化学合成法合成辅酶Q10, 将是工业化生产辅酶Q10的主要途径。
侧链直接引入法合成辅酶Q10的关键因素是:(1) 反应过程中聚异戊烯长侧链中双键几何构型的保持;(2) 防止反应过程中侧链副反应的发生;(3) 聚异戊烯长链与母体高收率的结合。直接进行Friedel-Crafts烷基化反应合成带短链的磺酰基化合物, 然后与茄尼基溴偶联反应生成辅酶Q10前体化合物, 大大简化了辅酶Q10前体的合成路线。侧链直接引入法是一条竞争力较为欠缺的化学合成路线, 辅酶Q10化学合成一直是较为活跃的研究领域。
以茄尼醇作原料进行辅酶Q10的合成是将茄尼醇经多步反应转化成多异戊烯基衍生物, 与带有氯甲基短支链的母环在镍催化下进行交叉偶联反应, 最后在钴催化下以DMF和PhMe作溶剂, 室温下反应24h, 在空气中氧化生成辅酶Q10, 收率64%, 改进了辅酶Q10的合成方法。新方法的最大改进之处是分别合成末端为碳铝功能基团的多异戊烯基衍生物和氯甲基辅酶Q10, 两者在镍催化下交叉偶联反应, 这是合成辅酶010的最后一步反应, 相对提高了茄尼醇的利用率, 经济价值高于以前的路线。
侧链延长法合成辅酶Q10的关键因素是:(1) 短侧链与母体化合物高效偶合;(2) 延长短侧链时双键几何构型的保持。该法克服了侧链直接引入法中将茄尼醇扩链为癸异戊烯基化合物时产生不易分离的顺反异构体混合物的不足, 从而提高了侧链构建时的偶合效率和立体选择性。最重要的是所用原料中最昂贵的茄尼醇, 得以在最终的交叉偶联步骤中引入整个合成路线, 大大提高了茄尼醇的利用率, 达到显著降低生产成本的目标, 因此寻求工艺优良的侧链延长法是化学合成辅酶Q10的发展方向。
其次需合成出具有立体选择性的多戊烯醇有卤化物(一般采用茄尼醇为出发原料) , 最后是进行两者高产率的缩合。无论是侧链引入还是侧链延长方式, 都需要对母核化合物进行基团保护, 在缩合时, 还需要考虑产物的构型问题。
侧链直接引入法合成辅酶Q10的3个关键问题是:(1) 反应过程中聚异戊二烯链中双键构型如何保持;(2) 防止反应过程中侧链副反应的发生;(3) 聚异戊二烯链部分与母核化合物高产率的结合。
侧链延长法合成辅酶Q10的关键问题是:(1) 短侧链与母体化合物高产率的偶合;(2) 延长短侧链时双键几何构型的保持。
8.1.3 香兰素法
以香兰素为原料合成TMB的反应过程是在0.25L压力釜中, 加入27%的甲醇钠甲醇溶液160g、乙酸乙酯40ml和干燥的5-溴香兰素46g, 常温搅拌25min。再加入催化剂氯化亚铜, 通入二氧化碳后进行密闭升温反应。控制温度135℃, 于压力0.65~0.70MPa下反应3h, 然后降温停止反应。将物料转移至蒸馏瓶中, 回收甲醇。加入水100ml, 加热使反应混合物充分溶解, 冷冻至5℃使产物结晶、吸滤, 得黄绿色丁香醛酚钠湿品。
丁香醛酚钠盐无需酸化、干燥, 可直接用于甲基化反应。取少许丁香醛酚钠盐经酸化、热滤、干燥后得丁香醛。
在上述所得的丁香醛酚钠盐湿品中加水及少量活性炭纤维, 加热到微沸保温30min脱色, 趁热滤除活性炭纤维和催化剂。滤液冷却到40℃, 滴加碳酸二甲酯和液碱进行甲基化反应。先滴入碳酸二甲酯后再缓慢滴加液碱调整pH在9.0, 待碱性不变时, 再滴入碳酸二甲酯, 以液碱调整pH在规定范围, 经多次滴完后, 再将液温升至50℃, 保温30min结束反应, 冷却至10℃。过滤, 以水洗至中性, 干燥, 得白色颗粒状固体TMB, 收率84%, 含量稳定在99%以上。此法避免了溶剂的精制回收过程, 节约能源, 减少污染, 降低生产成本, 改善了操作条件。
8.1.4 生物提取法
根据提取工艺的不同又分为:醇-碱皂化制造法和醇-醚混合提取法。
(1) 醇-碱皂化制造法
该工艺辅酶Q10的收率为60.9mg/kg新鲜猪心, 这是国内普遍采用的提取工艺。
在乙醇存在时, 长时间皂化可能导致辅酶Q10环上的甲氧基与乙醇中的乙氧基换位, 生成单或双乙氧基衍生物, 为避免杂质的生成, 可以使用KOH代替氢氧化钠和甲醇皂化。但必须加入焦性没石子酸, 否则辅酶Q10将在皂化过程中全部被破坏。加入量占起始湿原料的6%, 皂化过程中可通氮气。
(2) 醇-醚混合提取法
该工艺操作同醇-碱皂化制造法基本相似, 只是省去了皂化反应。
在上述两种方法中醇-碱皂化制造法对辅酶Q10的提纯较为迅速、完全, 且易于纯化, 但需要消耗大量的焦性没石子酸, 在皂化过程中产生乙氧基衍生物。醇-醚混合提取法的收率不如皂化法, 但可以省去焦性没石子酸, 不会产生乙氧基衍生物。
8.1.5 微生物发酵法
8.1.5.1 操作过程
1977年实现了微生物发酵法生产辅酶Q10。这种生产工艺被认为是最有前途的合成工艺, 近几年微生物发酵法成为国内外开发的热点。红极毛杆菌、烷微环菌等是生产辅酶Q10的主要菌种。
微生物发酵法是近些年发展起来的新的生物技术, 因为它能避免天然提取和化学合成法的缺点, 是很有发展产途的新技术。微生物法工业化生产主要条件是有稳定的规模化生产工艺和高产菌种, 同时要有现代的分离技术。
微生物发酵技术可在常温或稍高于常温与在常压下进行, 其特点是技术要求较高, 因发酵条件控制不好、菌种纯化保藏不善、生产原料批次与质量波动都会使发酵单位发生很大差距。此外, 微生物发酵技术还需一套复杂的质监测定方法。目前仍使用活性炭或大孔吸附剂吸附发酵清液后解吸、浓缩等常规后处理技术。产品纯度为80%。
在生物合成途径中, 几十倍地提高产率的关键是必须选育使用高效抗药菌种。
在辅酶Q10发酵生产中, 生产菌株的选择是非常重要的。人们广泛寻找辅酶Q10产生菌, 发现了许多高产菌株。
光合细菌菌体中辅酶Q10的含量普遍较高, 因此是产辅酶Q10菌种的理想选择之一。但野生型菌株产辅酶Q10能力不能满足生产需要, 这就需要利用常规诱变及基因工程技术对其进行遗传改造。辅酶Q10的微生物合成途径, 主要分为劈香环合成及异戊二烯基侧链的生物合成路线, 根据这两种生物合成途径再结合细菌的代谢调节机制, 我们就可以有目的的对产辅酶Q10菌株进行筛选。
由异戊烯基侧链及芳香环的生物合成途径路线, 选育营养缺陷型突变株, 有利于提高所需产物的浓度。类胡萝卜素与辅酶Q10均以聚异戊二烯为前体进行合成代谢, 减少类胡萝卜素的生成量可能会促进辅酶Q10的生物合成。
除了应用代谢控制理论选育高产突变株或构建重组菌株以提高发酵辅酶Q10产量外, 对生产菌的发酵条件进行优化也是提高发酵产量的一条重要而有效的途径。
菌种生长的优劣, 培养基的成分极其重要, 不同菌种, 培养基的成分也不同, 在辅酶Q10发酵中可通过选择不同来源成分的碳源、氮源、生长因子、无机盐等进行发酵优化实验, 以确定培养基的组成。有研究表明, 金属离子对发酵生产辅酶Q10有促进作用。另外, 在培养基中添加一定量的其他物质, 也能明显提高辅酶Q10的产量。在脱氮副球菌的基本培养基中加入胆碱可提高辅酶Q10的合成量约达30%;豆油、西红柿汁、烟叶、胡萝卜素、桔子皮汁的添加能大幅度提高酵母菌中辅酶Q10的含量, 微生物中辅酶Q10的合成与胡萝卜素合成密切相关。
培养模式分为分批发酵、流加发酵、补料、连续发酵等, 不同的培养模式各有其优缺点, 培养模式可直接影响辅酶Q10的生成量。目前, 采用补料分批培养更具前景。分批补料可以避免代谢———积累而得到较高的细胞密度, 延长发酵过程, 积累大量产物;通过辅酶Q10生产菌的呼吸强度、糖代谢与辅酶Q10生产的相关关系研究及氧供应量与葡萄糖供应的动力学分析, 发酵过程中葡萄糖维持在较低的浓度有利于产物的形成, 因此, 也可采用流加培养。对放射性根瘤菌研究发现, 采用蔗糖和适宜量玉米浆混合流加发酵能显著提高辅酶Q10产量, 其胞内辅酶Q10含量提高33%。
还可采用多级发酵系统, 多级发酵系统的不同阶段有恒温培养、通风培养等, 该系统对于缩短发酵时间, 节约生产费用, 提高产量等具有重要意义。
培养条件控制的好坏直接影响到代谢方式及代谢流量的变化, 从而影响到目的产物(次级代谢产物) 的产量, 了解培养条件对过程的影响, 可以帮助人们有效地控制微生物的生长和生产。
微生物的生长和代谢除了需要适宜的营养环境外, 其它环境因子也应处于适宜的状态, 其中pH是极为重要的一个环境因子。初始pH可影响菌体对基质的利用速度和细胞的结构状态, 以致影响菌体生长速度和代谢产物的变化。p H不同, 辅酶Q10的生成量也会有所不同, 对豌豆根瘤菌的研究发现, 该菌在pH 5.0~10.0范围内均能生长, 并且出现了2个最适生长pH, 分别为5.0和9.0;但在酸性条件下比碱性条件下生成的辅酶Q10质量高。这说明酸性条件更有利于辅酶Q10的生物合成。另一方面, 除考虑营养的需求外, 也要考虑其代谢后对培养体系pH缓冲体系的贡献, 从而保证整个发酵过程中pH能够处于较为适宜的状态。
微生物的生长和产物的合成均需在其各自适合的温度下进行, 而且温度是保证酶活性的重要条件, 故在发酵过程中必须保证最适宜的环境。一般发酵温度升高, 酶反应速率增大, 生长代谢加快, 生产期提前, 但酶易因过热而失去活性, 影响最终产量。对黄色隐球酵母的研究发现, 28~30℃下酵母生长量最优, 但辅酶Q10的产量高峰在26~28℃, 因此, 将温度设定为28~29℃, 酵母生长良好, 辅酶产量也较高。
通气对辅酶Q10产量的影响因生产菌株的不同而异, 一种情况是能促进辅酶Q10的发酵生产, 另一种情况是具有抑制作用。在多数情况下, 好氧发酵是多种微生物进行辅酶Q10发酵生产的基本特征。在一定供氧范围内, 其产物醌类含量随通气强度及搅拌转速的增加而增加。
影响辅酶Q10产量的培养因素还有接种量、搅拌转速、光照、培养时间等, 通过实验研究, 对其进行优化, 从而制定出一套培养条件适合的方案, 提高辅酶Q10的产量。
由于辅酶Q10为细胞内物质, 主要存在于细胞内线粒体内膜上, 其提取效果的好坏直接影响到产品的纯度和得率, 目前提取辅酶Q10的方法有不经皂化的提取法和皂化提取法。采用不经皂化的提取法提取辅酶Q10的量较少, 但不会破坏辅酶Q10;皂化提取法是经典提取脂溶性物质的方法, 方法虽简单, 但成本较高。最近提出的碱皂化提取法可以完全省去焦性没食子酸、乙醇溶剂的消耗费用, 而采用酸破碎细胞直接皂化, 使辅酶Q10的生产成本大幅度下降。采用碱皂化法从粟酒裂殖酵母中提取辅酶Q10, 优化后辅酶Q10产量比之前提高33.7%。
微生物发酵法是近些年发展起来的新的生物技术, 因为它能避免天然提取和化学合成法的缺点, 操作实例如下:
例1菌种的培养基及培养。
(1) 培养基
种子培养基(g/L) :葡萄糖20、淀粉40、磷酸二氢钾l、硫酸镁0.1、玉米浆50、VB10.01、pH为6.4;
发酵培养基(g/L) :淀粉40、植物油40、豆饼粉40、玉米浆30、磷酸二氢钾1、硫酸镁0.1、VB10.01, pH为6.4。
(2) 菌种培养
种子培养:取斜面上少量菌丝接入(装液量50ml/250ml) 种子培养基中, 置于27℃、210r/min旋转摇床上振荡培养42h;
发酵培养:将培养好的种子以15%(v/v) 的接种量转入发酵培养基中(装液量40ml/500ml) , 置于27℃、210r/min的旋转摇床上振荡培养72h。
(3) 湿菌丝体的制备
用纱布将菌丝体从发酵液中过滤出来, 去离子水洗涤两次, 拧干得湿菌丝体, 作为提取样品。称取0.5g湿菌丝体, 置于干燥箱中干燥, 恒重后称重, 并计算菌体的干湿比, 进而计算出菌丝体的干重。剩余湿菌体即为皂化提取的样品。
(4) 皂化法提取菌丝体中辅酶Q10
称取上述湿菌体2g, 移入250ml平底烧瓶内, 按湿菌体质量的l0%加入焦性没食子酸、19倍的100A KOH的甲醇溶液及少量去离子水混匀。在70℃水浴锅中回流60min后, 用自来水迅速冷却至室温, 倒入500ml分液漏斗中, 分3次加石油醚萃取辅酶Q10。合并萃取液, 用去离子水洗涤至中性, 加无水硫酸钠干燥。用旋转蒸发仪浓缩后移至10ml容量瓶中, 用石油醚定容至刻度, 待测定。
(5) 标准品的配制
精密称取辅酶‰标准品50mg, 置于10ml容量瓶中, 加1ml氯仿使之完全溶解, 再用无水乙醇定容至刻度, 制成5rng/ml的标准品溶液。
例2菌种为放射型根瘤菌放线菌素D和L乙基硫氨酸双抗性突变株Rhizobium radiobacter WSH-E25。
(1) 培养基
斜面和种子培养基(g/L) :葡萄糖20、蛋白胨10、酵母膏10、氯化钠5、pH为7.2、121℃灭菌15min。斜面培养基另加琼脂20g/L。
发酵培养基(g/L) :葡萄糖30、糖蜜60、酵母膏5、(NH4) 2SO47、玉米浆30、KH2PO40.4、Na2HPO4·12H2O 1.5, MgSO4·7H2O 0.4、pH为7.2、121℃灭菌15min。
(2) 菌种培养
斜面活化和种子培养:将冰箱中保藏的斜面菌种接种到活化斜面上, 30℃静置培养24h;将活化后斜面菌种接扩种子培养基, 30℃200r/min振荡培养24h。摇瓶装液量为50m/250ml三角瓶, 发酵培养。
取种子培养液以7%(v/v) 接种量接入发酵培养基, 装液量为50ml/500ml三角瓶, 30℃、200r/min振荡培养80h。
HPLC分析。
细胞干重测定:取发酵液4ml, 于10, 000r/min离心10min, 收集菌体, 用蒸馏水洗涤二次, 105℃烘干至恒重后测定。
例3菌种的培养基及培养。
(1) 斜面培养基
PDA培养基。种子培养基:波美麦芽汁7%、葡萄糖2%、酵母膏0.5%、pH自然;
发酵培养基:葡萄糖3%、酵母膏0.5%、蛋白胨0.2%、(NH4) 2SO40.1%、玉米粉0.1%、MgSO40.05%、K2HPO40.05%、KH2PO40.1%、pH自然。
(2) 菌悬液的制备
将热带假丝酵母斜面种接入种子培养液中, 30℃振荡培养24h, 使其处于对数生长期, 离心收集菌体, 用0.1mol/L pH为7.0的磷酸缓冲液洗涤3次, 并制备成细胞浓度为106个/ml的菌悬液;另取1ml稀释至106个/ml, 作为计算致死率的空白对照。
硫酸二乙酯-紫外复合诱变:分别取细胞浓度为106个/ml的菌悬液lml, 加入DES使其终浓度为1%, 分别处理20、30、40、50、60min;转至平皿中, 再置紫外灯下分别照射30、45、60、75、90s, 各取0.3ml涂布于PDA平板中, 30℃避光培养, 参照空白对照计算致死率。硫酸二乙酯-紫外复合诱变结果如下:
通过DES-UV复合诱变, 在复合诱变作用下, 致死率基本上都在80%以上, 其中以1%的DES处理40min, 再经紫外处理60s后, 突变率达到了2.71%。
将复合诱变作为高产菌筛选的最佳处理方式, 以1%的DES处理40min, 再经紫外线处理60s的复合剂量作为诱变的最佳剂量。
通过放线菌素D对突变茵进行广谱抗性初筛, 并以高浓度前体物质对羟基苯甲酸以及终产物结构(类似物维生素K) , 进行特异性筛选, 选育得到遗传稳定性良好的辅酶Q10高产突变株APV.12, 使得APV.12菌株辅酶Q10的发酵产量达到23.958mg/L, 比出发菌株提高了1.76倍。
该法能较大幅度地提高辅酶Q10的发酵产量, 从而解决微生物发酵法生产辅酶Q10菌种发酵产量低的工业化难题。
以根癌土壤杆菌WSHATl2为出发菌株, 采用传统的诱变育种与高效的推理选育方法相结合的方式, 针对辅酶Q10合成途径的特异性进行诱变处理。首先定向筛选出抗L-乙硫氨酸突变株, 然后以该突变株为出发菌株, 筛选得到L-乙硫氨酸(Eth) 维生素比(Ⅵu) 双抗性突变株, 以双抗性突变株为出发菌株, 进一步诱变得到X-gal利用能力提高的突变株, 以提高菌株积累辅酶Q10的能力。
例4菌种为三孢布拉氏霉Blakeslea trispora JSF4。
近年来, 目前多采用酵母属及细菌发酵制备辅酶Q10, 采用三孢布拉氏霉发酵制备辅酶Q10。
(1) 培养基
孢子斜面和平皿分离培养基基。PDA培养抗性培养基:PDA+L甲硫氨酸或D-酪氨酸或对氨基苯丙氨酸。
种子培养基(mg/10l) :淀粉4、葡萄糖2、玉米浆5、KH2PO40.1、MgSO40.01、VBl0.001、pH为6.4。
发酵培养基(mg/10l) :淀粉2、黄豆饼粉4、玉米浆3、KH2PO40.1、MgSO40.01、VB10.001、大豆油2.5、pH为6.7。
(2) 抗性菌株的选育
将诱变紫外线处理过的孢子悬液作一定的梯度稀释, 涂布于抗性培养基上, 27℃培养4~5d, 挑取单菌落接于斜面。
(3) 培养方法
将斜面上的少量菌丝接种于种子培养基(250ml烧瓶装量50ml) 、置于27℃、210r/min的旋转摇床上振荡42h。
(4) 摇瓶发酵
将培养好的种子以15mg/10l的接种量转入发酵培养基(500ml烧瓶装量40ml) 中, 置于27℃、220r/min的旋转摇床上振荡72h。
从发酵液中过滤出菌丝体, 移入250ml平底烧瓶中, 加抗氧化剂、甲醇、少量蒸馏水混匀, 90℃回流30min, 迅速冷却, 用石油醚萃取3次, 合并萃取液进行减压浓缩。采用薄层层析分离浓缩液中的辅酶Q10。
采用微生物发酵法生产辅酶Q10得到的产品为天然品, 生物活性好, 易被人体吸收;而且没有原材料的制约, 反应条件温和, 可通过规模放大提高生产能力, 是辅酶Q10生产最具潜力的生产方法。
微生物发酵法作为近些年发展起来的新的生物技术, 它能避免天然提取和化学合成法的缺点, 是很有发展产途的新技术。微生物法实现工业化生产的主要条件是有稳定的规模化生产工艺和高产菌种, 同时要有现代的分离技术。
发酵法已取得了相当程度的进展。如制备出富含辅酶Q10的菌液, 为酵母菌含量的10倍, 牛肝的100倍。另外, 由于国外已采用全新的分离技术设备, 产品纯度可达99%以上。
微生物发酵法具有原料来源丰富、反应条件温和、产品生物活性好等优点而成为最有发展潜力的生产方法。
例5菌种出发菌株为根癌:土壤杆菌WSHATl2。
(1) 培养基
种子培养基(g/L) :葡萄糖10、酵母膏5、蛋白胨5、氯化钠5、pH为7.2、121℃灭菌l5min。斜面培养基另加琼脂20g/L。
平板培养基(g/L) :葡萄糖5, 酵母膏5, 蛋白胨10、MgSO4·H2O 2、琼脂20、pH为7.2、121℃灭菌15min。
含X-gal的基本培养基(g/L) :X-gal 0.2、葡萄糖100、硫酸铵5、KHSO40.4、NaHPO41.5、MgSO4·7H2O0.3、麦芽汁20、pH为7.2、121℃灭菌151min。
优化前发酵培养基(g/L) :葡萄糖20、酵母膏10、蛋白胨10、MgSO4·7H2O 0.5、Na2PO41.5、KH2PO40.5、pH为7.2、121灭菌15min。
优化后发酵培养基(g/L) :蔗糖30、玉米浆40、硫酸铵5、MgSPO4·7H2O 0.3, Na2HPO41、KH2PO40.6、pH为7.5、121C灭菌15min。
(2) 菌种培养方法
斜面活化和种子培养:将冰箱中保存的斜面菌种接种到新鲜斜面上活化, 30℃静置培养24h;将活化后的斜面菌种接人种子培养基, 在温度为30℃下以200r/min转速振荡培养24h。摇瓶培养实验装液量如无特指均为250ml。
(3) 发酵培养
取种子培养液以6%接种量接人发酵培养基, 在30℃温度下以200r/min转速振荡培养110h(除特别说明外) 。每个样品做3个平行样。
(4) 辅酶Q10提取
取3ml发酵液离心分离(10000r/min, 10min) 倾去上清液菌体于70℃冷冻24h加入3ml丙酮超声波振荡0.5h离心(10000r/min, 5min)
保存丙酮HPLC分析。
(5) 细胞干重(DCW) 的测定
取发酵液(3ml) 经10000r/min离心10min后倾去上清液, 去离子水洗2次, 菌泥经105℃干燥24h至恒重后称重。
(6) 诱变方法
将出发菌接种于种子培养基, 培养24h, 取5ml培养液10000r/min离心收集菌体, 用磷酸钾缓冲液(pH 7.0) 洗涤菌体2次, 制成浓度为107个/L的菌悬液。取10ml菌悬液至一无菌碘量瓶内, 加入酒精数滴后, 加入硫酸二乙酯(体积分数为2%) , 振荡15min, 加入5m l5%的硫代硫酸钠, 振荡10min, 吸取3ml于5ml离心管中离心收集菌体, 用磷酸钾缓冲液洗涤3次后, 接入种子培养基中培养12h后, 离心洗涤3次, 吸0.1ml稀释后直接涂布于含L-乙硫氨酸的培养基平板上, 于30℃培养4d, 挑选菌落直径比较大的单菌落于斜面培养。
X-gal利用能力提高的突变株的筛选:诱变后的菌液稀释后直接涂布于X-gal平板上, 培养4d。出发菌株的菌落为淡蓝色, 挑出呈深蓝色的菌落, 即为X-gaP突变株。
8.1.5.2 工艺特点
微生物发酵法作为近些年发展起来的新的生物技术, 它能避免天然提取和化学合成法的缺点, 是很有发展产途的新技术。微生物法实现工业化生产主要条件是有稳定的规模化生产工艺和高产菌种, 同时要有现代的分离技术。
发酵法已取得了相当程度的进展。如制备出富含辅酶Q10的菌液, 为酵母菌含量的10倍, 牛肝的100倍。采用全新的分离技术设备, 产品纯度可达99%以上。
微生物发酵法因具有原料来源丰富、反应条件温和、产品生物活性好等优点而成为最有发展潜力的生产方法。微生物发酵技术可在常温或稍高于常温与在常压下进行, 特点是技术要求较高, 因发酵条件控制不好, 菌种纯化保藏不善, 生产原料批次与质量波动都会使发酵单位发生很大差距。此外, 微生物发酵技术还需一套复杂的质监测定方法。目前仍使用活性炭或大孔吸附剂吸附发酵清液后解吸、浓缩等常规后处理技术。产品纯度为80%。
采用生物工程的方法, 从培养的微生物中提取的辅酶Q10具有生物活性高, 价格低的优势, 比进口辅酶Q10的价格便宜10倍左右, 有很好的市场竞争力。
采用微生物发酵法生产辅酶Q10得到的产品为天然品, 生物活性好, 易被人体吸收;而且没有原材料的制约, 可通过规模放大提高生产能力, 是辅酶Q10生产最具潜力的生产方法。
九、总结与展望
辅酶Q10具有肯定的抗氧化与抗疲劳作用。虽然临床研究的例数有限, 但其结果表明, 该成份可以作为中老年人的抗氧化剂用于多种疾病的辅助治疗, 并可有效缓解疲劳、改善氧化应激反应等, 其使用的安全性也很好。
辅酶Q10具有抗肿瘤、抗氧化性, 以及作为细胞代谢激活剂, 可提高人体免疫力和治疗人体免疫系统疾病, 特别在心血管疾病治疗中有重要作用。因此, 辅酶Q10可作为一种新型的具重大医学价值的生化药物或开发保健食品的良好材料。但是关于辅酶Q10在治疗临床疾病过程中的具体作用机理还不是很清楚, 尚需要进一步的研究。此外, 辅酶Q10与相关药物联合使用治疗疾病的效果会更好, 而且大量的研究也表明联合应用抗氧化剂可以产生协同或相加效应, 但目前此方面的研究不是很多, 提示未来可以开展更多种类更广范围的辅酶Q10与药物或抗氧化剂的结合来治疗相关疾病和产品的开发。
辅酶Q10预治疗可以增强卵巢对刺激的反应,改善卵巢储备下降的患者的卵母细胞和胚胎质量。对于DOR患者,给予一段时间的辅酶Q10预治疗对其卵巢功能恢复和未来妊娠有明显效果。已经证明,每天摄入高达900 mg的辅酶Q10对健康成年人是安全的。辅酶Q10的治疗将延迟辅助生殖技术周期,预治疗时长要根据患者情况而定,长期的预治疗患者依从性可能较差。由于年龄问题,有些患者不具备长期辅酶Q10预治疗的条件。需要进一步的实验研究来确定补充辅酶Q10最佳剂量和最佳持续时间以改善DOR。
由于辅酶Q10具有易溶于脂而难溶于水的物理性质, 因此, 在药物和化妆品方面有广泛的应用, 目前在食品领域, 辅酶Q10全面推广使用的困难是成本问题。辅酶Q10利用的关键是其生物利用度, 如何提高辅酶Q10的生物利用度是当前研究领域的焦点。使用固体分散体技术、环糊精包合技术、乳化技术、自微乳化技术及脂质体技术均能不同程度地改善辅酶Q10的水溶性和增强贮存过程中辅酶Q10的稳定性, 可提高其生物利用度, 丰富辅酶Q10的用途。随着人们对辅酶Q10的不断研究和深度开发, 它必将更好地为人类所用。
随着人们对辅酶Q10功能认识的不断深入和生活水平的不断提高, 可以预言, 未来辅酶Q10的市场前景将非常广阔。在科研方面, 关于辅酶Q10生物合成途径已经基本清晰, 但对于其代谢调节规律却依然知之甚少。因此, 在基础研究领域, 未来关于辅酶Q10的代谢调节将是研究的难点和重点。另外, 对高产菌种的选育或构建, 将直接决定辅酶Q10的生产率和成本。综合利用各种遗传手段, 构建出高水平的生产菌种, 将是应用研究领域的研究重点和难点。未来, 辅酶Q10的研究重点一方面是构建高产菌种、优化工艺条件从而降低生产成本;另一方面需要进一步研究代谢机制与应用方法, 使辅酶Q10的应用更方便。
